

Lateral organization of the postsynaptic density
- PMID: 21920440
- PMCID: PMC3216044
- DOI: 10.1016/j.mcn.2011.09.001
Lateral organization of the postsynaptic density
Abstract
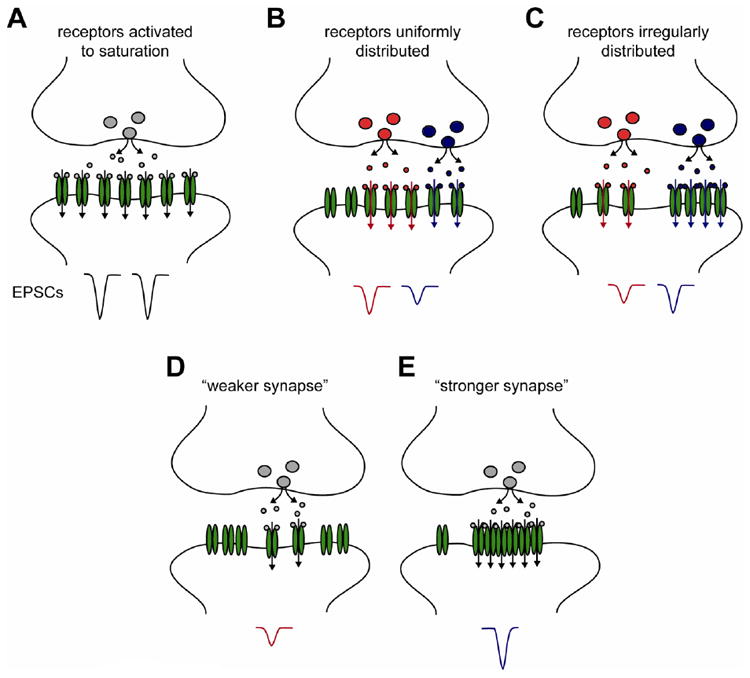
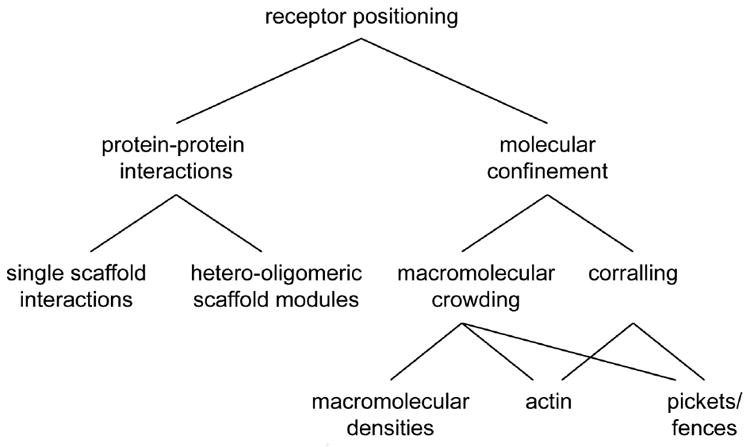
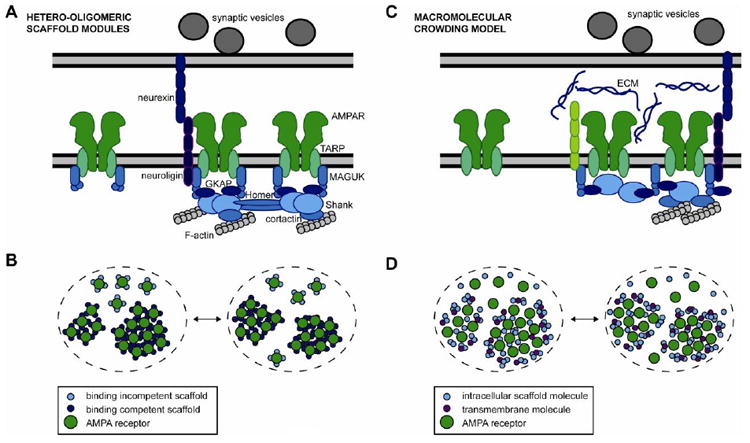
Fast excitatory synaptic transmission is mediated by AMPA-type glutamate receptors (AMPARs). It is widely accepted that the number of AMPARs in the postsynaptic density (PSD) critically determines the efficiency of synaptic transmission, but an unappreciated aspect of synapse organization is the lateral positioning of AMPARs within the PSD, that is, their distribution across the face of a single synapse. Receptor lateral positioning is important in a number of processes, most notably because alignment with presynaptic release sites heavily influences the probability of receptor activation. In this review, we summarize current understanding of the mechanisms that dynamically control the subsynaptic positioning of AMPARs. This field is still at early stages, but the recent wave of developments in super-resolution microscopy, synapse tomography, and computational modeling now enable the study of lateral protein distribution and dynamics within the nanometer-scale boundaries of the PSD. We discuss data available measuring the lateral distribution of glutamate receptors and scaffold proteins within the PSD, and discuss potential mechanisms that might give rise to these patterns. Elucidating the mechanisms that underlie the lateral organization of the PSD will be critical to improve our understanding of synaptic processes whose disruption may be unexpectedly important in neurological disorders. This article is part of a Special Issue entitled Membrane Trafficking and Cytoskeletal Dynamics in 'Neuronal Function'.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Adam G, Matus A. Role of actin in the organisation of brain postsynaptic densities. Brain Res Mol Brain Res. 1996;43:246–250. - PubMed
-
- Alber F, Dokudovskaya S, Veenhoff LM, Zhang W, Kipper J, Devos D, Suprapto A, Karni-Schmidt O, Williams R, Chait BT, et al. Determining the architectures of macromolecular assemblies. Nature. 2007;450:683–694. - PubMed
-
- Allen JA, Halverson-Tamboli RA, Rasenick MM. Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci. 2007;8:128–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
