

Ixodes scapularis salivary gland protein P11 facilitates migration of Anaplasma phagocytophilum from the tick gut to salivary glands
- PMID: 21921936
- PMCID: PMC3207102
- DOI: 10.1038/embor.2011.177
Ixodes scapularis salivary gland protein P11 facilitates migration of Anaplasma phagocytophilum from the tick gut to salivary glands
Abstract
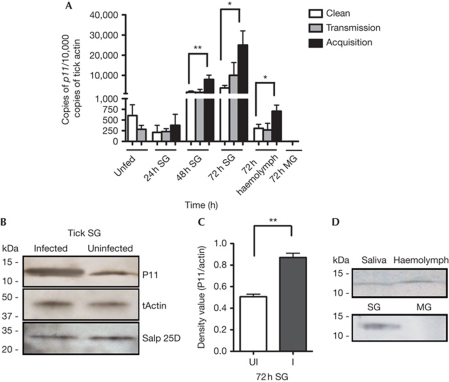
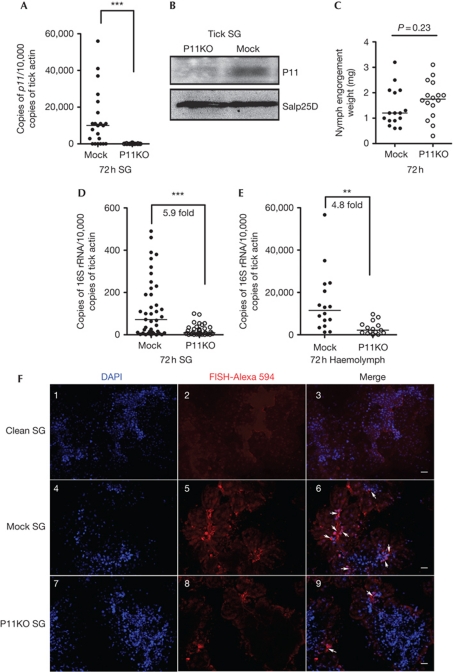
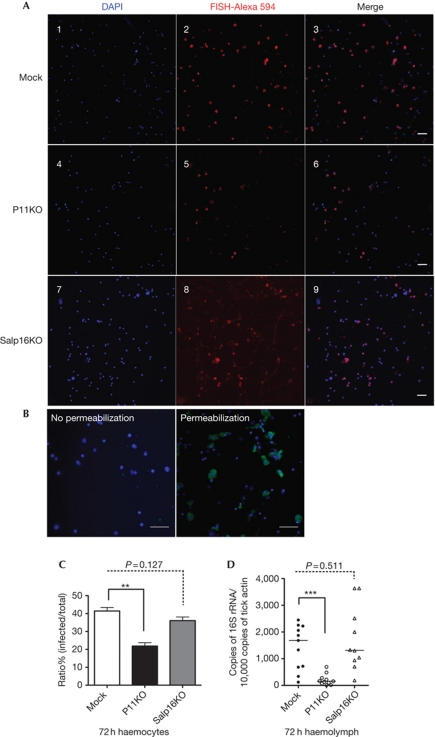
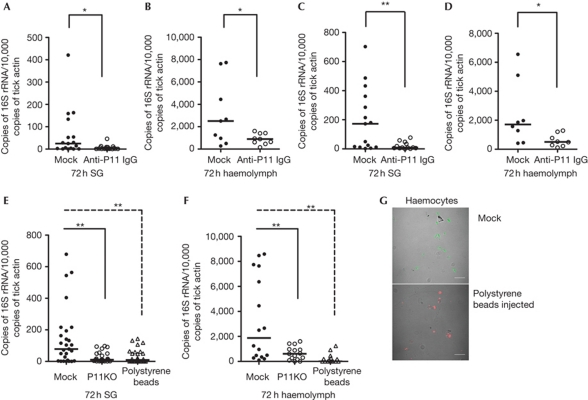
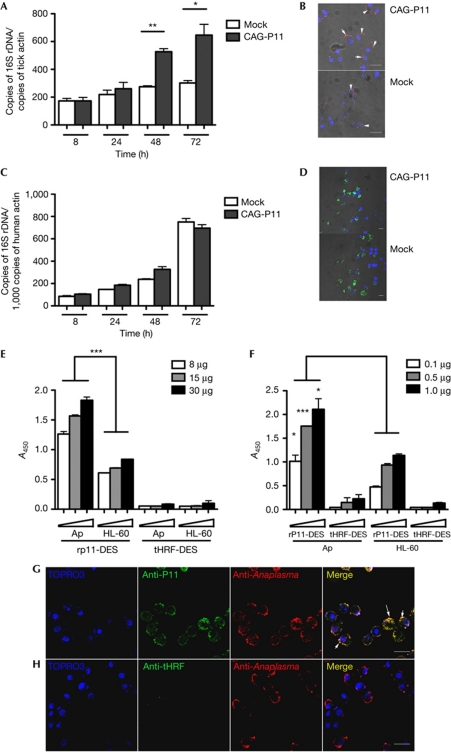
Ixodes ticks harbour several human pathogens belonging to the order Rickettsiales, including Anaplasma phagocytophilum, the agent of human anaplasmosis. When ticks feed on A. phagocytophilum-infected mice, the pathogen enters the ticks' gut. The bacteria then migrate from the gut to infect the salivary glands of the ticks and are transmitted to the next host via the saliva. The molecular mechanisms that enable the migration of A. phagocytophilum from the gut to the salivary glands are poorly understood. Here we show that a secreted tick protein, P11, is important in this process. We show that P11 enables A. phagocytophilum to infect tick haemocytes, which are required for the migration of A. phagocytophilum from the gut to the salivary glands. Silencing of p11 impaired the A. phagocytophilum infection of tick haemocytes in vivo and consequently decreased pathogen infection of the salivary glands. In vitro experiments showed that P11 could bind to A. phagocytophilum and thus facilitate its infection of tick cells. This report provides new insights into A. phagocytophilum infection of ticks and reveals new avenues to interrupt the life cycle of Anaplasma and related Rickettsial pathogens.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Bakken JS, Dumler JS (2000) Human granulocytic ehrlichiosis. Clin Infect Dis 31: 554–560 - PubMed
-
- Bakken JS, Dumler JS, Chen SM, Eckman MR, Van Etta LL, Walker DH (1994) Human granulocytic ehrlichiosis in the upper Midwest United States. A new species emerging? JAMA 272: 212–218 - PubMed
-
- Borovickova B, Hypsa V (2005) Ontogeny of tick hemocytes: a comparative analysis of Ixodes ricinus and Ornithodoros moubata. Exp Appl Acarol 35: 317–333 - PubMed
-
- Carlyon JA, Fikrig E (2006) Mechanisms of evasion of neutrophil killing by Anaplasma phagocytophilum. Curr Opin Hematol 13: 28–33 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
