

Functional roles of alpha-band phase synchronization in local and large-scale cortical networks
- PMID: 21922012
- PMCID: PMC3166799
- DOI: 10.3389/fpsyg.2011.00204
Functional roles of alpha-band phase synchronization in local and large-scale cortical networks
Abstract
Alpha-frequency band (8-14 Hz) oscillations are among the most salient phenomena in human electroencephalography (EEG) recordings and yet their functional roles have remained unclear. Much of research on alpha oscillations in human EEG has focused on peri-stimulus amplitude dynamics, which phenomenologically support an idea of alpha oscillations being negatively correlated with local cortical excitability and having a role in the suppression of task-irrelevant neuronal processing. This kind of an inhibitory role for alpha oscillations is also supported by several functional magnetic resonance imaging and trans-cranial magnetic stimulation studies. Nevertheless, investigations of local and inter-areal alpha phase dynamics suggest that the alpha-frequency band rhythmicity may play a role also in active task-relevant neuronal processing. These data imply that inter-areal alpha phase synchronization could support attentional, executive, and contextual functions. In this review, we outline evidence supporting different views on the roles of alpha oscillations in cortical networks and unresolved issues that should be addressed to resolve or reconcile these apparently contrasting hypotheses.
Keywords: alpha; amplitude; electroencephalography; magnetoencephalography; phase; source modeling; synchrony.
Figures
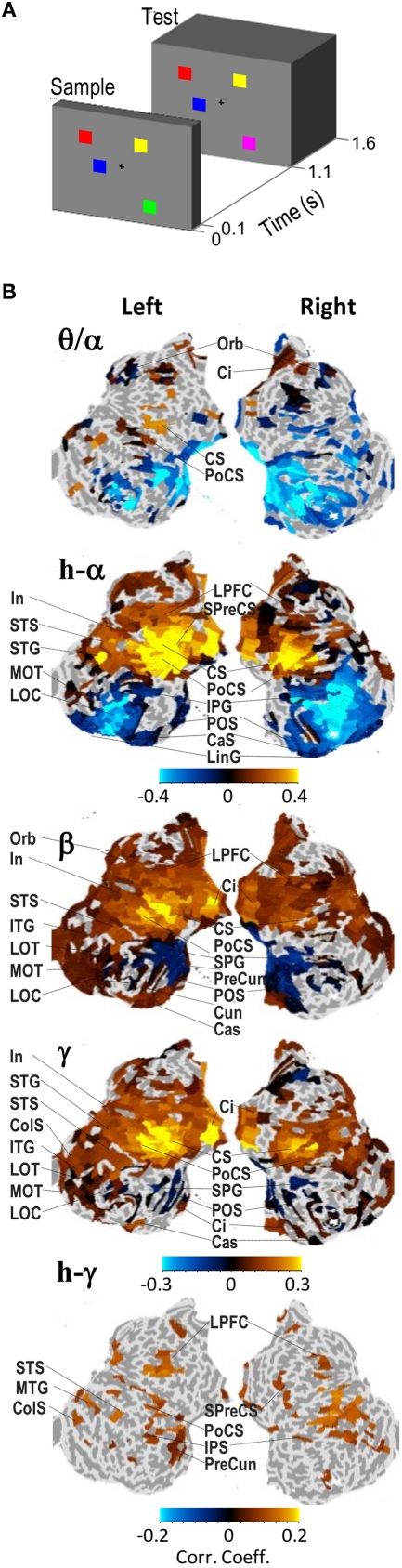
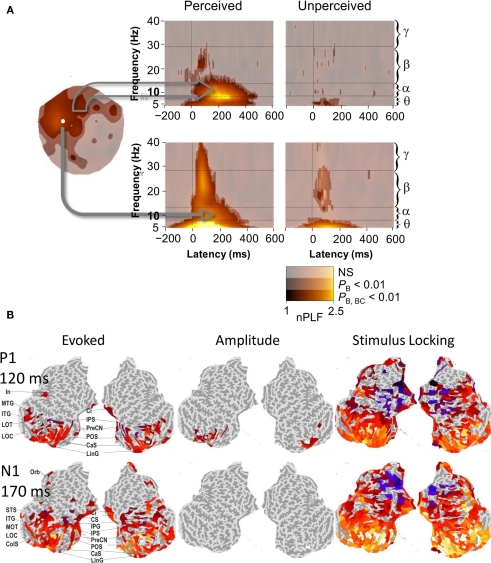
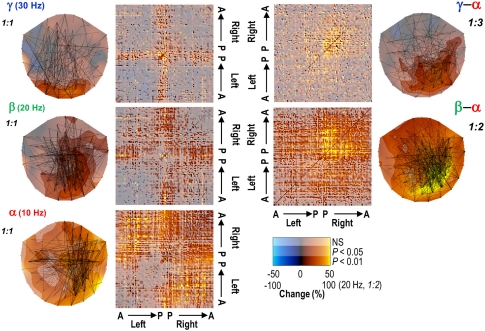
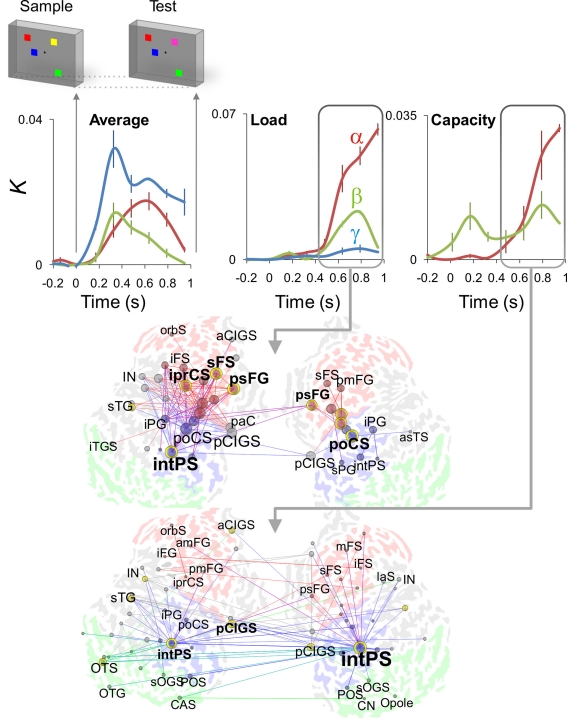
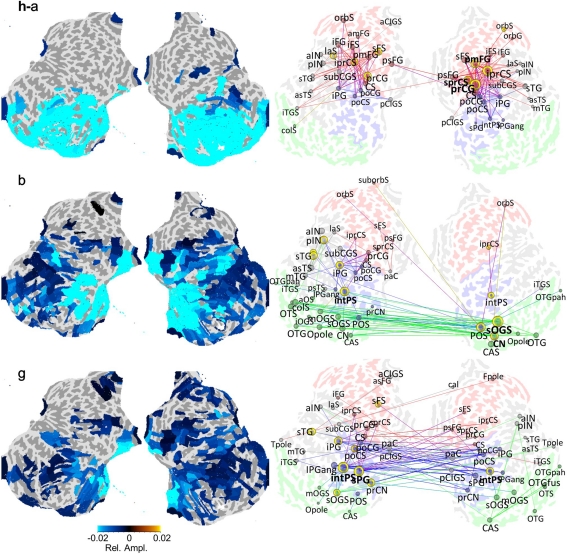
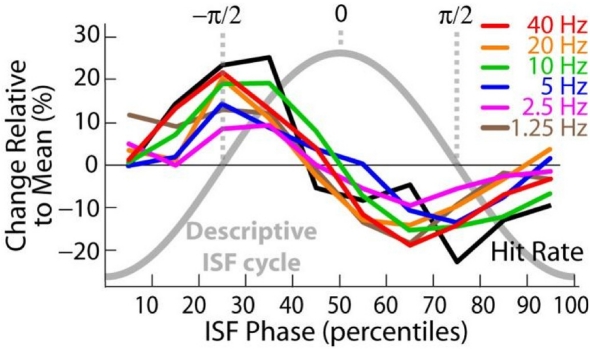
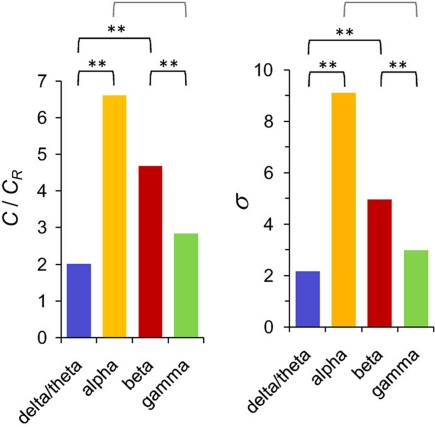
References
-
- Andersen E. A. S. (1968). Physiological Basis of the Alpha Rhythm New York, NY: Appleton-Century-Crofts
LinkOut - more resources
Full Text Sources