

Review
doi: 10.1021/cr2002799.
Epub 2011 Sep 19.
Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: cross-talk between the eicosanoid and endocannabinoid signaling pathways
Affiliations
- PMID: 21923193
- PMCID: PMC3191732
- DOI: 10.1021/cr2002799
Item in Clipboard
Review
Endocannabinoid oxygenation by cyclooxygenases, lipoxygenases, and cytochromes P450: cross-talk between the eicosanoid and endocannabinoid signaling pathways
Chem Rev.
.
Free PMC article
No abstract available
Figures
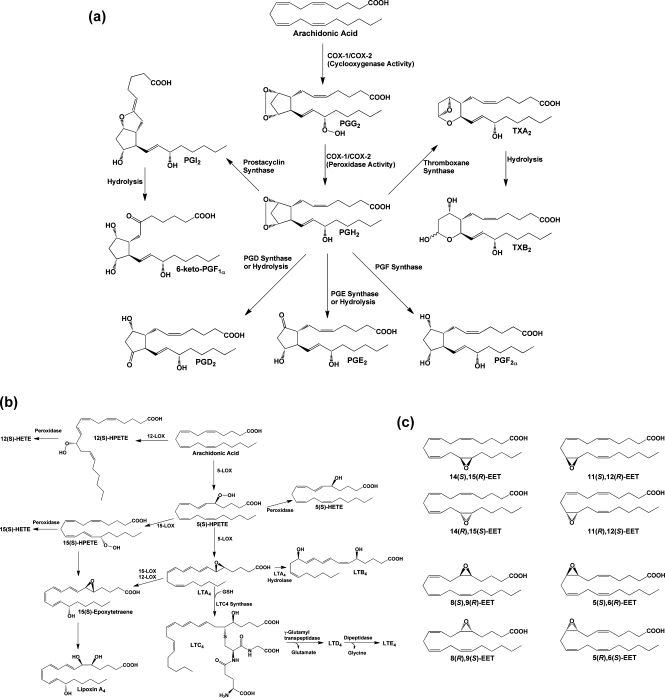
(a) Cyclooxygenase pathway of AA metabolism. AA is converted to PGG2 at the cyclooxygenase active site of COX-1 or COX-2 and is then reduced to PGH2 at the peroxidase active site. PGH2 spontaneously decomposes to yield PGE2 or PGD2, but these compounds, as well as PGF2α, PGI2, and TxA2, are also produced enzymatically by specific synthases. PGI2 and TXA2 are unstable and spontaneously decompose to yield 6-keto-PGF1α and TXB2, respectively. The ethanolamide and glyceryl ester of PGH2 produced from the metabolism of AEA and 2-AG, respectively, are converted to the same range of eicosanoid products as PGH2 with the exception of the TXA2 analogue. (b) Lipoxygenase pathway of AA metabolism. AA is converted by LOX enzymes to position-specific HPETEs, which are then reduced to the corresponding HETEs. 5-LOX also converts 5-HPETE to LTA4, which may then be metabolized to LTB4 or LTC4. The glutathionyl moiety of LTC4 is subject to enzymatic hydrolysis, yielding LTD4 and LTE4. The actions of multiple LOX enzymes lead to the formation of lipoxins. An example is provided for the synthesis of lipoxin A4 through multiple steps catalyzed by 5-LOX and either 15-LOX or 12-LOX. (c) Cytochromes P450 catalyze the epoxygenation of AA at each of the double bonds, producing the range of products shown.
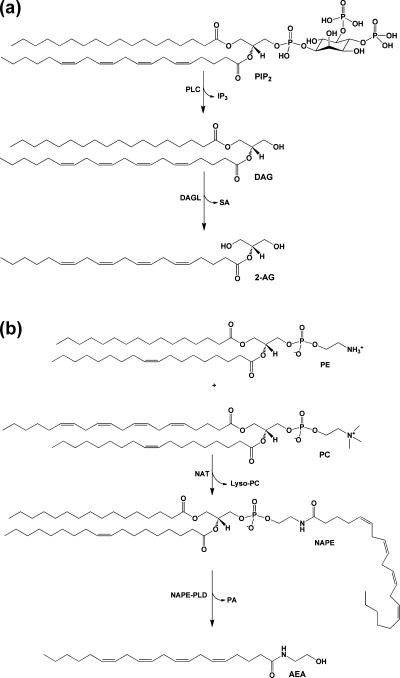
(a) Biosynthetic pathway for 2-AG. PIP2 containing AA at the sn-2 position is hydrolyzed by PLC to yield DAG and IP3. DAG is then further hydrolyzed by MAG lipase to 2-AG. (b) Biosynthetic pathway for AEA. An arachidonoyl moiety is transferred from the sn-1 position of a phospholipid, in this case PC, to the amino group of PE by NAT. The resulting product NAPE is then hydrolyzed by NAPE-PLD to yield AEA and PA.
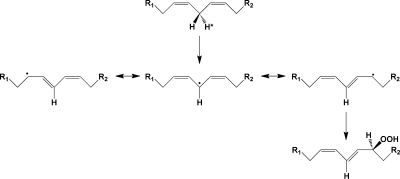
Mechanism of the lipoxygenase reaction. A hydrogen atom is abstracted from the bisallylic carbon of a polyunsaturated fatty acid, yielding a 1,5-pentadienyl radical. Addition of oxygen at the terminus of this radical yields a peroxyl radical, which is then reduced to the hydroperoxide.
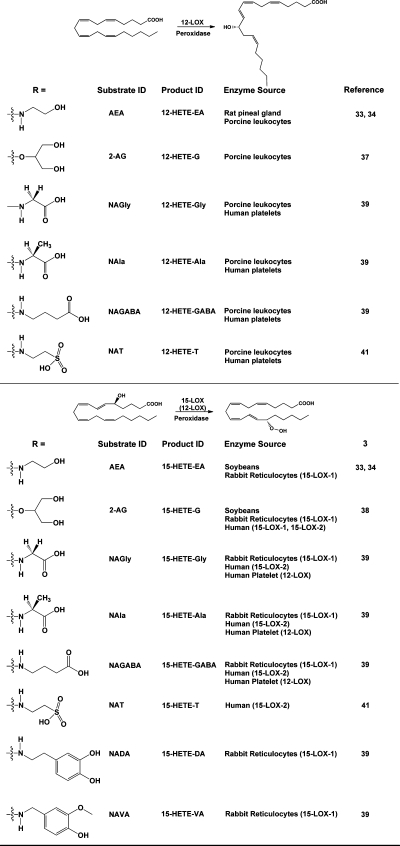
Products of the action of lipoxygenases on endocannabinoids and related AA-derived compounds. The general reactions of 12-LOX and 15-LOX are shown, along with the structures of the endocannabinoid analogues that have been tested as substrates. The identities of the major products are indicated, along with the sources of the enzymes that have been shown to catalyze the reaction. Note that, in the case of the human platelet 12-LOX metabolizing arachidonoyl amino acids, both 12- and 15-hydroxylated products were identified. For this reason, 12-LOX is shown (in parentheses) as catalyzing the formation of this positional isomer.
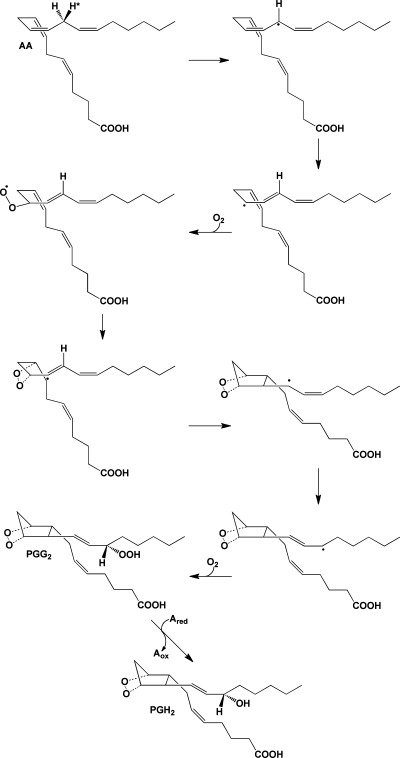
Mechanism of the cyclooxygenase reaction. The 13-pro-(S)-hydrogen of AA is removed by a radical at Y385 of the COX active site. The resulting radical migrates to position 11, which serves as the site of oxygen addition. Following the formation of the endoperoxide between carbons 11 and 9, a single bond links carbons 8 and 12 to form the prostanoid five-membered ring. The radical then migrates to carbon 15, which becomes the site of the second oxygen addition, forming a peroxyl radical, which is then reduced to a hydroperoxide (PGG2). Reduction of the 15-hydroperoxide using electrons from a coreductant (Ared) at the peroxidase active site yields PGH2.
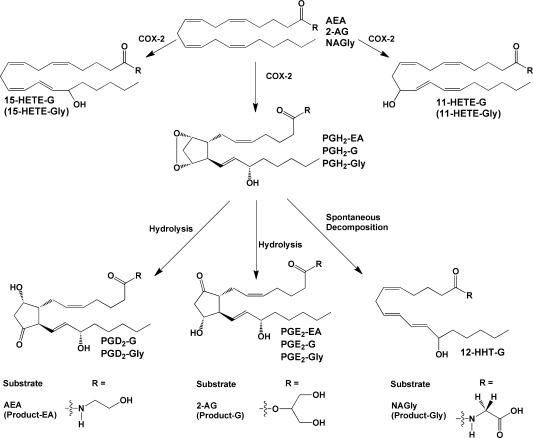
Products of the action of cyclooxygenase on endocannabinoids and related compounds. COX-2 produces PGH2 as the primary product, plus 11- and 15-HETE as minor products. In the absence of any additional metabolizing enzymes, PGH2 decomposes to form PGD2, PGE2, and HHT. Some or all of these compounds have been identified as products of the metabolism of endocannabinoid-like compounds as indicated above. For NAGly, a HETE-Gly derivative(s) was detected, but the exact position(s) of the hydroxyl group(s) was not determined. Endocannabinoid metabolism (2-AG and AEA) has been demonstrated for human COX-2; however, recombinant murine COX-2 has been used in the majority of these studies.
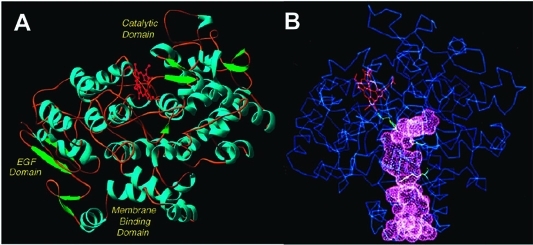
(A) Domain structure of the COX enzymes. The N-terminus (not visible in the crystal structure) connects to the epidermal growth factor (EGF) domain. The EGF domain, in turn, connects to the membrane-binding domain, which comprises four α-helices (A–D). Helix D connects the membrane-binding domain to the large catalytic domain. α-helices are shown in blue, β-sheets are shown in green, and the heme prosthetic group is shown in red. (B) Drawing of the COX structure highlighting the hydrophobic channel of the cyclooxygenase active site. The large lobby region, which opens into the membrane-binding domain, is separated from the L-shaped channel above by a constriction. Key catalytic residues Tyr-385, Arg-120, and Ser-530 are shown in green. The heme prosthetic group is in red. These structural characteristics shown here for ovine COX-1 are representative of both isoforms. Panel B kindly provided by M. Garavito. Both figures reprinted from ref (44). Copyright 2003 American Chemical Society.
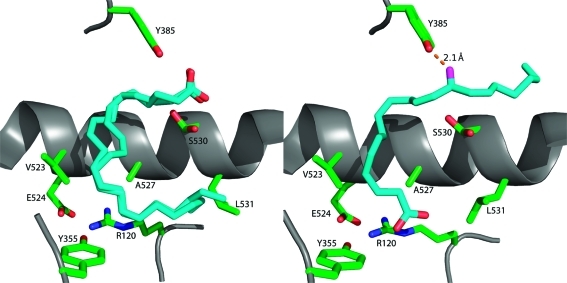
Comparison of the conformations of AA in the active sites of the two monomers of murine COX-2. The left frame displays AA bound in monomer A of the COX-2 homodimer. It is bound in an inverted catalytically unproductive conformation in which its carboxyl group is H-bonded to Tyr-385 and Ser-530. The ω end of the fatty acid lies across Arg-120 and abuts Leu-531. Two molecules of AA are modeled in monomer A because of ambiguities in modeling from the electron density map. The right frame displays AA bound in monomer B of the homodimer in a productive conformation. The carboxylate is H-bonded to Arg-120, and the fatty acid chain projects up into the active site. The ω end of the fatty acid projects into an alcove above Ser-530, and the 13-pro-(S) hydrogen is located adjacent to the hydroxyl group of Tyr-385, which is converted to a tyrosyl radical during turnover.
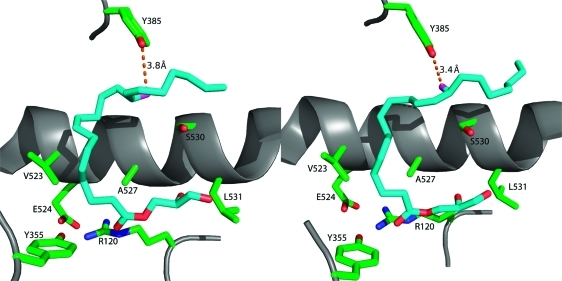
Comparison of the conformations of 1-AG in the active sites of the two monomers of murine COX-2. The left frame displays 1-AG in an unproductive conformation in monomer A of the homodimer, whereas the right frame displays 1-AG bound in a productive conformation in monomer B of the homodimer. The two conformations are comparable, but the ω ends of the fatty acyl groups differ in conformation so that the 13-pro-(S) hydrogen is only close enough to Tyr-385 in monomer B to enable abstraction during the catalytic cycle.
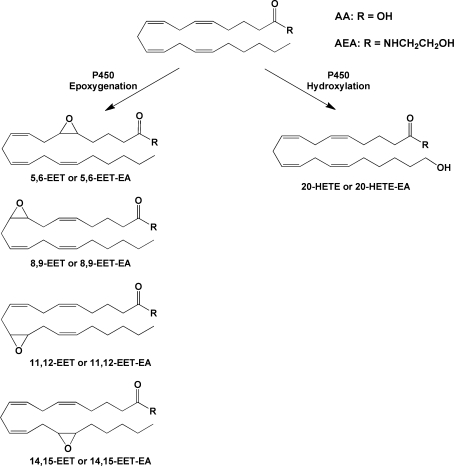
Examples of P450-catalyzed oxygenations of AA or AEA. Reactions shown are those that have been identified for AEA as well as AA. Additional reactions for AA have been reported.
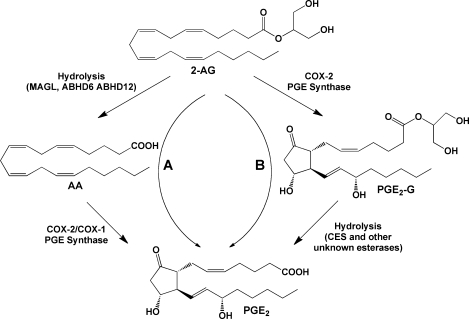
Hydrolytic metabolism of 2-AG and PG-Gs. Pathway A: 2-AG may be hydrolyzed to AA, which is then subject to oxygenation by COX-2 or COX-1, yielding free acid PGs (illustrated here by PGE2). Pathway B: Oxygenation of 2-AG will produce PG-Gs (illustrated here by PGE2-G), hydrolysis of which will yield the corresponding free acid PG. The origin of the free acid PG product, through oxygenation of AA or 2-AG, cannot readily be distinguished.
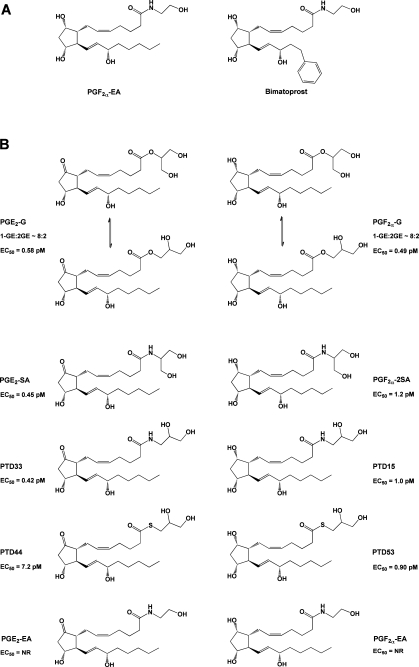
(A) Structures of PGF2α-EA and bimatoprost. (B) Structures of compounds tested for the ability to mobilize Ca2+ in RAW264.7 cells and H1819 cells.(115) Note that, in aqueous solution, PG-Gs rapidly equilibrate to form a mixture of the 1- and 2-glyceryl esters, with the 1-glyceryl ester predominating at a ratio of approximately 8:2. In contrast, the stereochemistry of the corresponding amide analogues remains fixed. EC50 values are given for Ca2+ mobilization in H1819 cells. NR indicates no response.
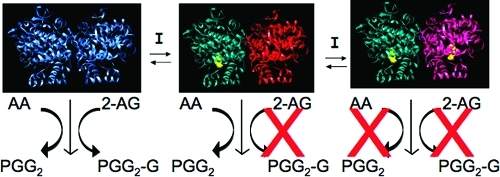
Mechanism of substrate-selective inhibition of endocannabinoid oxygenation by rapid, reversible inhibitors of COX-2. Inhibitor binding in one subunit of the homodimer induces a conformational change in the second subunit, which blocks 2-AG and AEA oxygenation but not AA oxygenation. Another molecule of inhibitor must bind in the second subunit to inhibit AA oxygenation. Reprinted from ref (134). Copyright 2009 American Chemical Society.
Similar articles
-
Oxidative metabolism of endocannabinoids.Prostaglandins Leukot Essent Fatty Acids. 2002 Feb-Mar;66(2-3):211-20. doi: 10.1054/plef.2001.0359. Prostaglandins Leukot Essent Fatty Acids. 2002. PMID: 12052037 Review.
-
Endocannabinoids and their oxygenation by cyclo-oxygenases, lipoxygenases and other oxygenases.Biochim Biophys Acta. 2015 Apr;1851(4):366-76. doi: 10.1016/j.bbalip.2014.12.015. Epub 2014 Dec 24. Biochim Biophys Acta. 2015. PMID: 25543004 Review.
-
Non-redundant functions of cyclooxygenases: oxygenation of endocannabinoids.J Biol Chem. 2008 Mar 28;283(13):8065-9. doi: 10.1074/jbc.R800005200. Epub 2008 Feb 4. J Biol Chem. 2008. PMID: 18250160 Free PMC article. Review.
-
Oxygenation of Anandamide by Lipoxygenases.Methods Mol Biol. 2023;2576:307-316. doi: 10.1007/978-1-0716-2728-0_26. Methods Mol Biol. 2023. PMID: 36152198
-
Eicosanoids: generation and detection in mammalian cells.Methods Mol Biol. 2009;462:5-23. Methods Mol Biol. 2009. PMID: 19160658 Review.
Cited by
-
Pharmacognosy and Effects of Cannabinoids in the Vascular System.ACS Pharmacol Transl Sci. 2022 Oct 28;5(11):1034-1049. doi: 10.1021/acsptsci.2c00141. eCollection 2022 Nov 11. ACS Pharmacol Transl Sci. 2022. PMID: 36407955 Free PMC article. Review.
-
Monoacylglycerol lipase inhibitors: modulators for lipid metabolism in cancer malignancy, neurological and metabolic disorders.Acta Pharm Sin B. 2020 Apr;10(4):582-602. doi: 10.1016/j.apsb.2019.10.006. Epub 2019 Oct 18. Acta Pharm Sin B. 2020. PMID: 32322464 Free PMC article. Review.
-
Fluorescent indomethacin-dansyl conjugates utilize the membrane-binding domain of cyclooxygenase-2 to block the opening to the active site.J Biol Chem. 2019 May 31;294(22):8690-8698. doi: 10.1074/jbc.RA119.007405. Epub 2019 Apr 18. J Biol Chem. 2019. PMID: 31000626 Free PMC article.
-
The Endocannabinoid System as a Therapeutic Target in Glaucoma.Neural Plast. 2016;2016:9364091. doi: 10.1155/2016/9364091. Epub 2016 Jan 12. Neural Plast. 2016. PMID: 26881140 Free PMC article. Review.
-
Plant-Derived and Endogenous Cannabinoids in Epilepsy.Clin Drug Investig. 2016 May;36(5):331-40. doi: 10.1007/s40261-016-0379-x. Clin Drug Investig. 2016. PMID: 26892745 Review.
References
-
- Devane W. A.; Dysarz F. A. 3rd; Johnson M. R.; Melvin L. S.; Howlett A. C. Mol. Pharmacol. 1988, 34, 605. - PubMed
-
- Matsuda L. A.; Lolait S. J.; Brownstein M. J.; Young A. C.; Bonner T. I. Nature 1990, 346, 561. - PubMed
-
- Munro S.; Thomas K. L.; Abu-Shaar M. Nature 1993, 365, 61. - PubMed
-
- Devane W. A.; Hanus L.; Breuer A.; Pertwee R. G.; Stevenson L. A.; Griffin G.; Gibson D.; Mandelbaum A.; Etinger A.; Mechoulam R. Science 1992, 258, 1946. - PubMed
-
- Sugiura T.; Kondo S.; Sukagawa A.; Nakane S.; Shinoda A.; Itoh K.; Yamashita A.; Waku K. Biochem. Biophys. Res. Commun. 1995, 215, 89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
