

Review
doi: 10.1016/j.bbamcr.2011.08.017.
Epub 2011 Sep 8.
Structure and function of the membrane deformation AAA ATPase Vps4
Affiliations
- PMID: 21925211
- PMCID: PMC3245771
- DOI: 10.1016/j.bbamcr.2011.08.017
Item in Clipboard
Review
Structure and function of the membrane deformation AAA ATPase Vps4
Biochim Biophys Acta.
2012 Jan.
Abstract
The ATPase Vps4 belongs to the type-I AAA family of proteins. Vps4 functions together with a group of proteins referred to as ESCRTs in membrane deformation and fission events. These cellular functions include vesicle formation at the endosome, cytokinesis and viral budding. The highly dynamic quaternary structure of Vps4 and its interactions with a network of regulators and co-factors has made the analysis of this ATPase challenging. Nevertheless, recent advances in the understanding of the cell biology of Vps4 together with structural information and in vitro studies are guiding mechanistic models of this ATPase.
Published by Elsevier B.V.
Figures
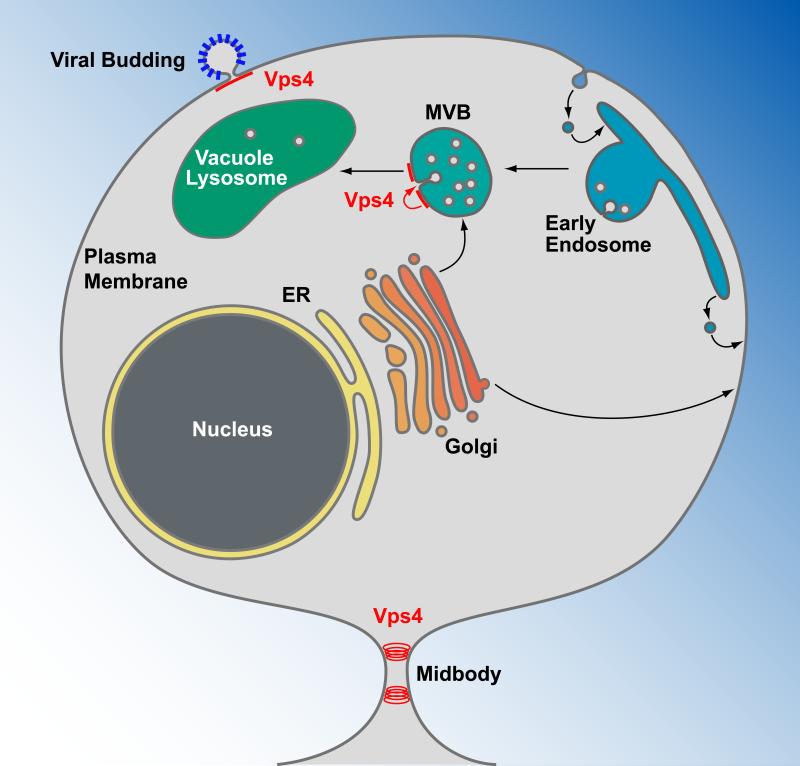
Vps4 functions in the formation of MVB vesicles, viral budding and cytokinesis. Transmembrane proteins are synthesized at the endoplasmic reticulum (ER), transported through the Golgi network and delivered either to the plasma membrane or an endosomal compartment. Plasma membrane proteins destined for degradation are endocytosed and delivered to endosomes. At the multivesicular body (MVB), these proteins are sorted by Vps4 and the ESCRTs into MVB vesicles (red arrow). Upon fusion of the MVB with the vacuole/lysosome, the lumenal vesicles are exposed to a hydrolytic environment, causing the degradation of proteins and lipids of the vesicles. Vps4 together with a subset of the ESCRTs are recruited to the plasma membrane by newly forming enveloped viruses, such as HIV-1, where they function in the release of the virus particles. At the final stage of cytokinesis, Vps4 is recruited to the midbody where it functions together with ESCRT-III in the abscission of the membrane and the formation of two separate cells.
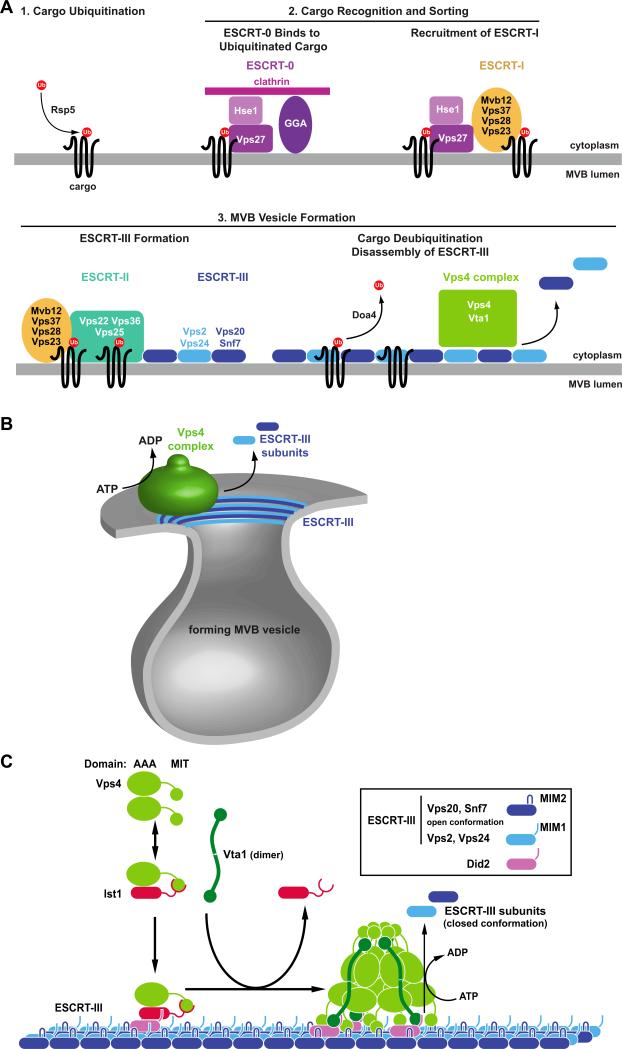
Models for the function of Vps4 and the ESCRT protein complexes. (A) Epistasis model for the ubiquitin-dependent sorting of transmembrane proteins (cargo) into the MVB pathway. Flat clathrin together with GGA proteins and ESCRT-0 (Vps27-Hse1) form a protein network at the endosome that binds ubiquitinated cargo. Vps27 recruits ESCRT-I to the MVB, where it supports the cargo sorting function. ESCRT-I interacts with ESCRT-II, resulting in the formation of ESCRT-III. ESCRT-I, ESCRT-II and ESCRT-III are involved in the formation of MVB vesicles that contain the cargo proteins that have been deubiquitinated by Doa4. The disassembly of ESCRT-III by Vps4 drives abscission of the vesicle neck and recycles the ESCRT factors for further rounds of sorting. (B) Model for the function of the Vps4/ESCRT-III system in the abscission of the vesicle neck. ATP-driven disassembly of ESCRT-III by Vps4 constricts the ESCRT-III collar at the neck of the forming vesicle, which induces membrane fission and vesicle formation. (C) Model for the recruitment and assembly of Vps4 on ESCRT-III. Ist1 binds to Vps4, which, together with Did2, aids the recruitment of Vps4 to MVB-associated ESCRT-III. Vta1 dimers support the assembly of Vps4 into the active double-ring structure. The fully assembled Vps4 complex hydrolyses ATP in order to disassemble ESCRT-III.
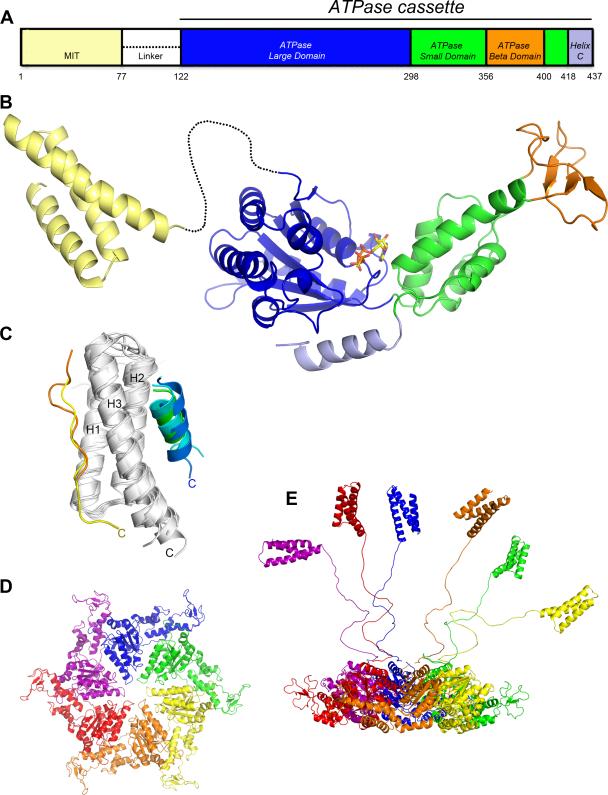
Structure of Vps4. (A) Schematic of the domain organization. (B) Composite ribbon diagram of the MIT domain and ATPase cassette. ATP-γ-S is bound at the ATP binding site between the large and small ATPase domains. (C) Overlap of independently determined structures of Vps4 MIT domains alone [45][47][48] and in complex with MIM1 peptides [48] blue, [47] green, [47] cyan or MIM2 peptides [24] yellow, [39] orange peptides. MIM1 and MIM2 peptides lie parallel to Helix 3 (C-termini are labeled), which places in them in an orientation to extend from the ESCRT-III complex and potentially bind the same MIT domain. (D) Model for the Vps4 hexamer based on the p97 D1 structure [49]. Top view showing only the ATPase cassettes. (E) Side view including MIT domains and linkers in a variety of potential relatively extended conformations. The extent to which linkers and MIT domains interact with the ATPase cassette hexamer is not currently defined.
References
-
- Odorizzi G, Babst M, Emr SD. Fab1p PtdIns(3)P 5-kinase function essential for protein sorting in the multivesicular body. Cell. 1998;95:847–858. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
