

Cone photoreceptor contributions to noise and correlations in the retinal output
- PMID: 21926983
- PMCID: PMC3183110
- DOI: 10.1038/nn.2927
Cone photoreceptor contributions to noise and correlations in the retinal output
Abstract
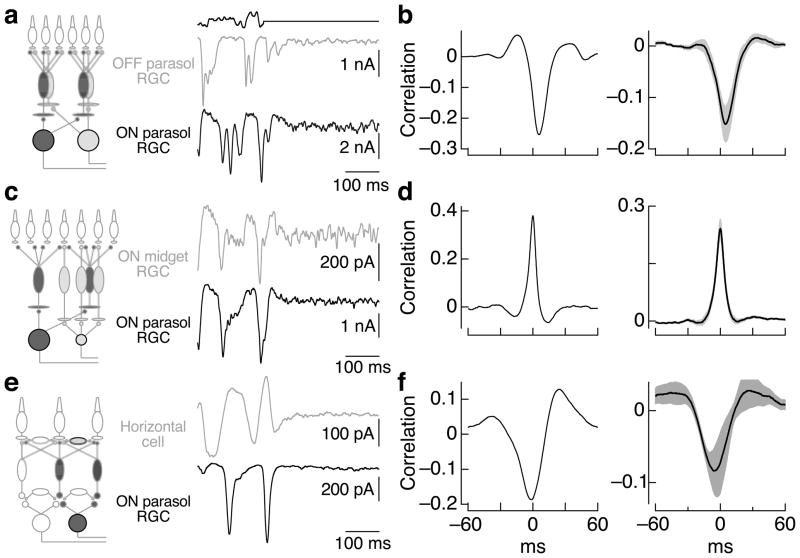
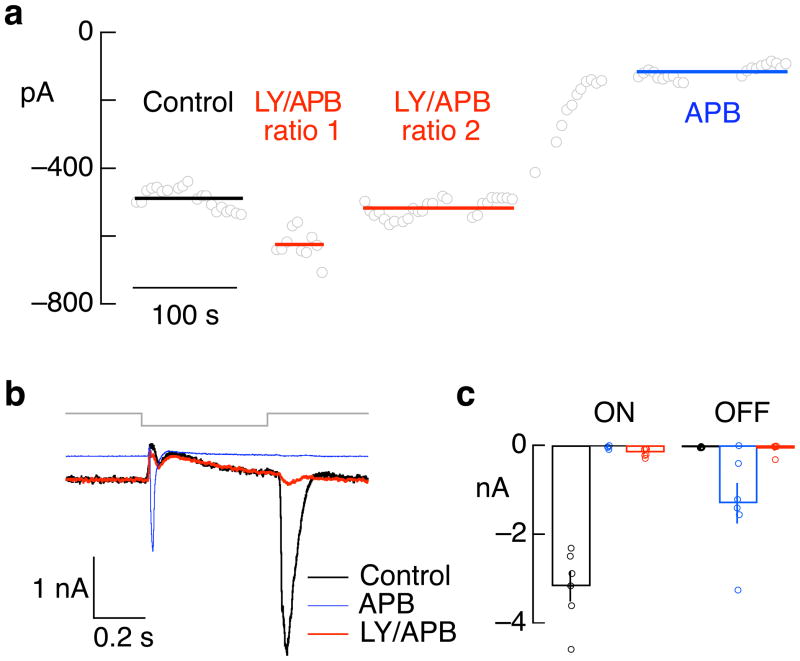
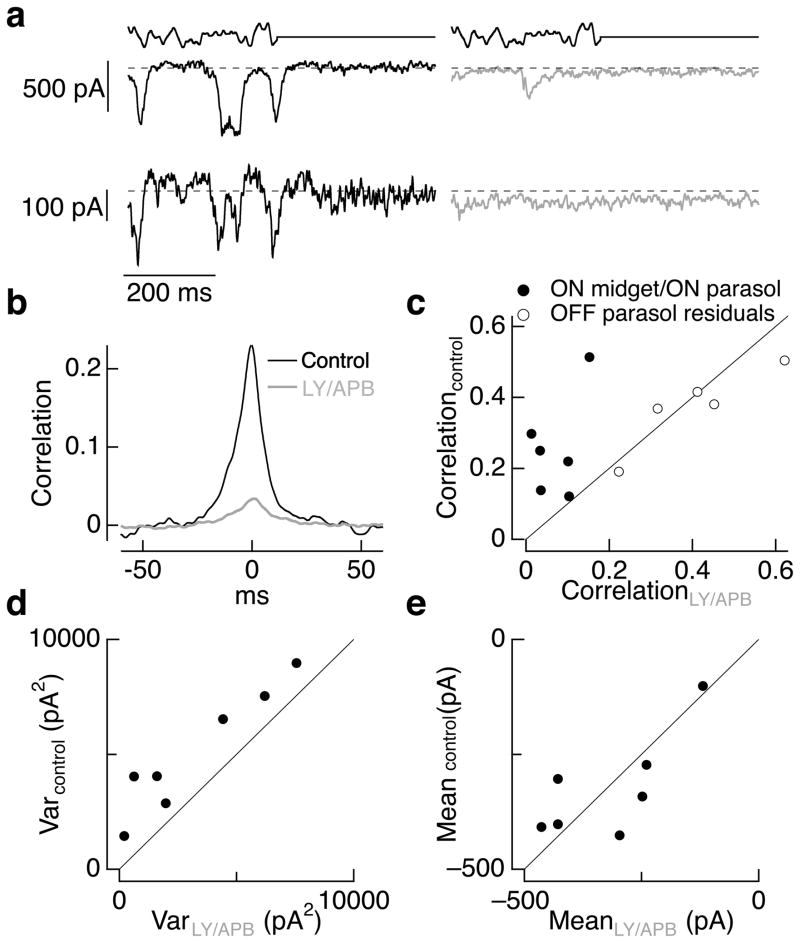
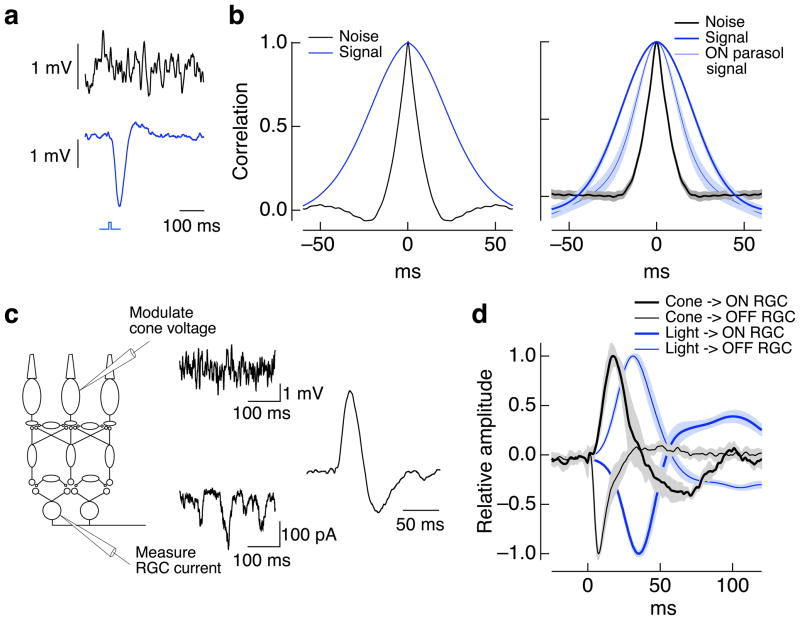
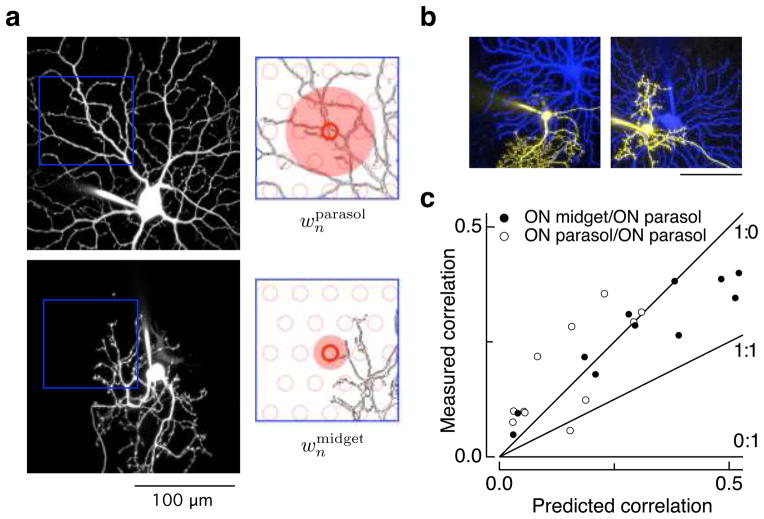
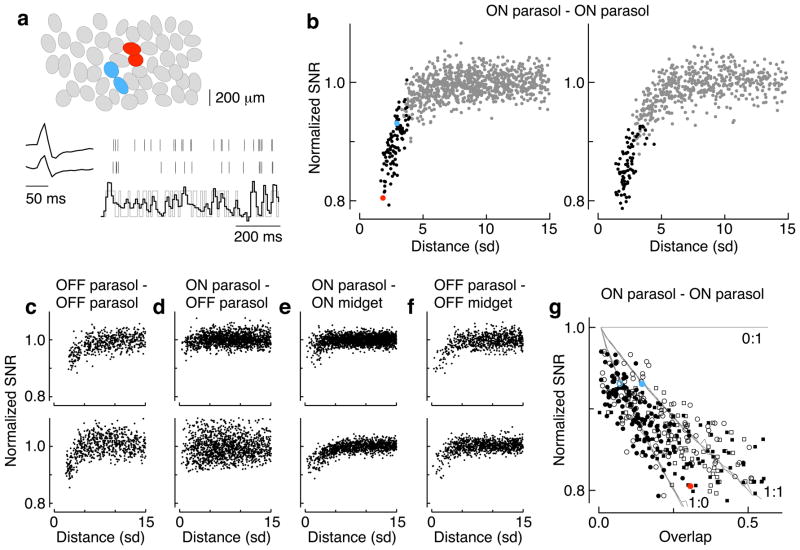
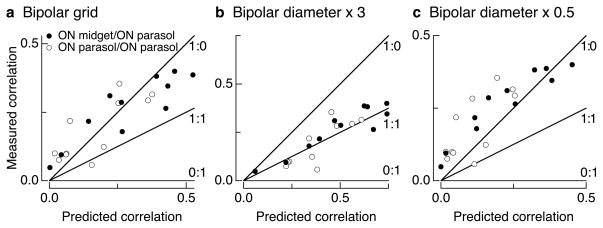
Transduction and synaptic noise generated in retinal cone photoreceptors determine the fidelity with which light inputs are encoded, and the readout of cone signals by downstream circuits determines whether this fidelity is used for vision. We examined the effect of cone noise on visual signals by measuring its contribution to correlated noise in primate retinal ganglion cells. Correlated noise was strong in the responses of dissimilar cell types with shared cone inputs. The dynamics of cone noise could account for rapid correlations in ganglion cell activity, and the extent of shared cone input could explain correlation strength. Furthermore, correlated noise limited the fidelity with which visual signals were encoded by populations of ganglion cells. Thus, a simple picture emerges: cone noise, traversing the retina through diverse pathways, accounts for most of the noise and correlations in the retinal output and constrains how higher centers exploit signals carried by parallel visual pathways.
© 2011 Nature America, Inc. All rights reserved.
Figures
References
-
- Mollon JD, Astell S, Cavonius CR. A reduction in stimulus duration can improve wavelength discriminations mediated by short-wave cones. Vision Res. 1992;32:745–755. - PubMed
-
- Westheimer G. Visual hyperacuity. Prog Sens Physiol. 1981;1:1–30.
-
- BARLOW HB. Purkinje shift and retinal noise. Nature. 1957;179:255–256. - PubMed
-
- Donner K. Noise and the absolute thresholds of cone and rod vision. Vision Res. 1992;32:853–866. - PubMed
-
- Barlow HB. Retinal noise and absolute threshold. J Opt Soc Am. 1956;46:634–639. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
