

doi: 10.1038/nchembio.659.
Metabolomics annotates ABHD3 as a physiologic regulator of medium-chain phospholipids
Affiliations
- PMID: 21926997
- PMCID: PMC3201731
- DOI: 10.1038/nchembio.659
Item in Clipboard
Metabolomics annotates ABHD3 as a physiologic regulator of medium-chain phospholipids
Nat Chem Biol.
.
Abstract
All organisms, including humans, possess a huge number of uncharacterized enzymes. Here we describe a general cell-based screen for enzyme substrate discovery by untargeted metabolomics and its application to identify the protein α/β-hydrolase domain-containing 3 (ABHD3) as a lipase that selectively cleaves medium-chain and oxidatively truncated phospholipids. Abhd3(-/-) mice possess elevated myristoyl (C14)-phospholipids, including the bioactive lipid C14-lysophosphatidylcholine, confirming the physiological relevance of our substrate assignments.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
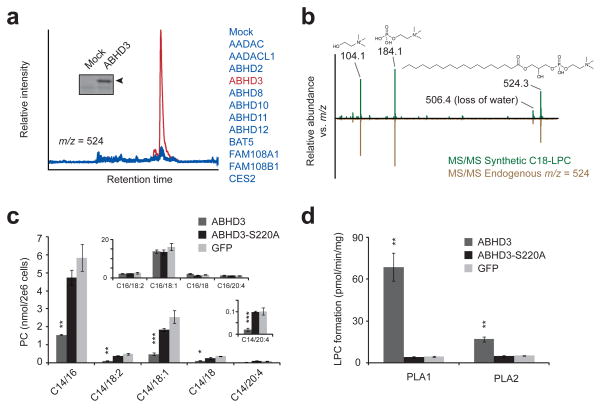
Metabolomic profiling of an enzyme library identifies metabolites altered by ABHD3 overexpression. (a) Representative overlaid extracted ion chromatograms at m/z = 523.5–524.5 from HEK293T cells transfected with ABHD3 (red trace) versus other enzymes (blue traces). Insert: Activity-based labeling using the serine hydrolase-directed probe fluorophosphonate rhodamine (FP-Rh) in mock versus ABHD3-transfected cells. Arrowhead designates FP-Rh-labeled ABHD3 protein. (b) Representative MS/MS fragmentation of synthetic C18-LPC (top, in green) and endogenous m/z = 524 (bottom, in brown) gave identical daughter ions of 104.1 (choline), 184.1 (phosphocholine), and 506.1 (dehydro-C18-LPC). (c) Targeted MRM measurements of phosphocholines (PCs) from C8161 cells stably overexpressing epitope-tagged ABHD3 (dark grey), the catalytically dead ABHD3-S220A mutant (black), or GFP (light grey). PC species are indicated by C#/##, where the # indicates the sn-1 acyl chain and ## indicates the sn-2 acyl chain. Top insert: C16-containing PC species; bottom insert: an enlarged graph showing C14/20:4-PC. (d) PLA1 and PLA2 hydrolysis activities for ABHD3 using C14/18:2-PC as a substrate and lysates from stably transfected C8161 cells as the protein source. Data are presented as mean ± standard error; n = 4/group; * P < 0.05, ** P < 0.01, *** P < 0.001 for ABHD3 versus ABHD3-S220A groups.
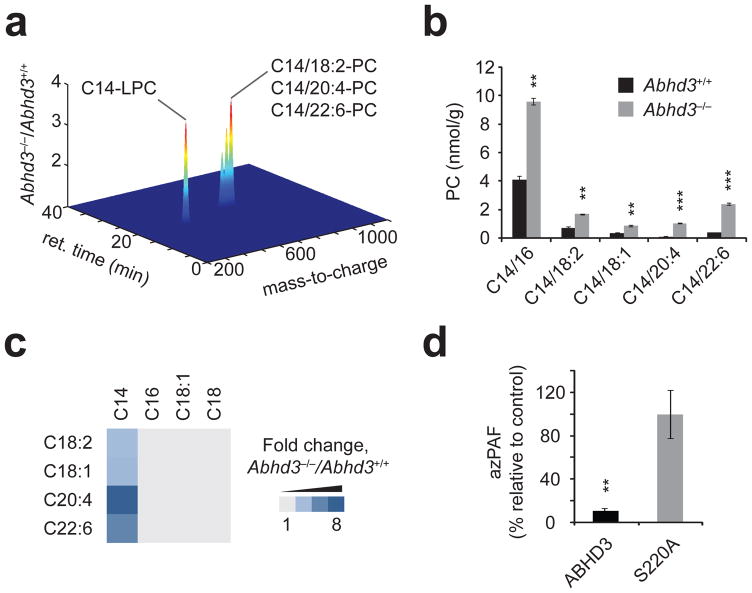
Abhd3−/− mice possess elevated C14-PCs. (a) Untargeted LC-MS profiling of kidney metabolomes from Abhd3+/+ or Abhd3−/− mice (plot is shown in the positive ionization mode). Only those metabolites with P < 0.05 and fold change > 1.5 are shown. Data are presented as mean fold changes; n = 6/group. (b) Targeted MRM measurements of PCs from kidney tissue of Abhd3+/+ (black bars) or Abhd3−/− (grey bars) mice. Similar changes in PCs were observed in brain, liver, and plasma from Abhd3−/− mice (see Supplementary Table 3). (c) Fold change of PC species from kidney tissue of Abhd3−/− versus Abhd3+/+ mice. The PC acyl chains for a given species are indicated by the top and left axis. Lighty grey indicates no change between genotypes whereas the darkest blue indicates a 8.5-fold change. Data are presented as means; n = 4/group. (d) Relative levels of azPAF in C8161 cells following live-cell incubation with azPAF (10 μM, 4 h, 37°C). For (b) and (d), data are presented as mean ± standard error; n = 3–4/group; * P < 0.05, ** P < 0.01, *** P < 0.001 for Abhd3+/+ versus Abhd3−/− or ABHD3 versus ABHD3-S220A.
References
-
- Lander ES. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921. - PubMed
-
- Gerlt JA, Babbitt PC. Divergent evolution of enzymatic function: mechanistically diverse superfamilies and functionally distinct suprafamilies. Annu Rev Biochem. 2001;70:209–46. - PubMed
-
- Raamsdonk LM, et al. A functional genomics strategy that uses metabolome data to reveal the phenotype of silent mutations. Nature Biotechnology. 2001;19:45–50. - PubMed
Publication types
MeSH terms
Substances
Associated data
- PubChem-Substance/125092044
- PubChem-Substance/125092045
- PubChem-Substance/125092046
- PubChem-Substance/125092047
- PubChem-Substance/125092048
- PubChem-Substance/125092049
- PubChem-Substance/125092050
- PubChem-Substance/125092051
- PubChem-Substance/125092052
- PubChem-Substance/125092053
- PubChem-Substance/125092054
- PubChem-Substance/125092055
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
