

Mechanical strain in actin networks regulates FilGAP and integrin binding to filamin A
- PMID: 21926999
- PMCID: PMC3204864
- DOI: 10.1038/nature10430
Mechanical strain in actin networks regulates FilGAP and integrin binding to filamin A
Abstract
Mechanical stresses elicit cellular reactions mediated by chemical signals. Defective responses to forces underlie human medical disorders such as cardiac failure and pulmonary injury. The actin cytoskeleton's connectivity enables it to transmit forces rapidly over large distances, implicating it in these physiological and pathological responses. Despite detailed knowledge of the cytoskeletal structure, the specific molecular switches that convert mechanical stimuli into chemical signals have remained elusive. Here we identify the actin-binding protein filamin A (FLNA) as a central mechanotransduction element of the cytoskeleton. We reconstituted a minimal system consisting of actin filaments, FLNA and two FLNA-binding partners: the cytoplasmic tail of β-integrin, and FilGAP. Integrins form an essential mechanical linkage between extracellular and intracellular environments, with β-integrin tails connecting to the actin cytoskeleton by binding directly to filamin. FilGAP is an FLNA-binding GTPase-activating protein specific for RAC, which in vivo regulates cell spreading and bleb formation. Using fluorescence loss after photoconversion, a novel, high-speed alternative to fluorescence recovery after photobleaching, we demonstrate that both externally imposed bulk shear and myosin-II-driven forces differentially regulate the binding of these partners to FLNA. Consistent with structural predictions, strain increases β-integrin binding to FLNA, whereas it causes FilGAP to dissociate from FLNA, providing a direct and specific molecular basis for cellular mechanotransduction. These results identify a molecular mechanotransduction element within the actin cytoskeleton, revealing that mechanical strain of key proteins regulates the binding of signalling molecules.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
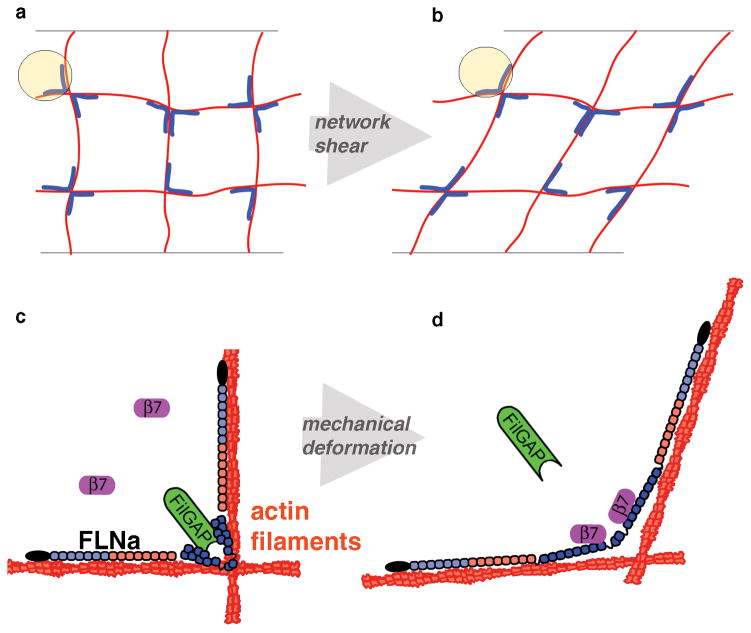
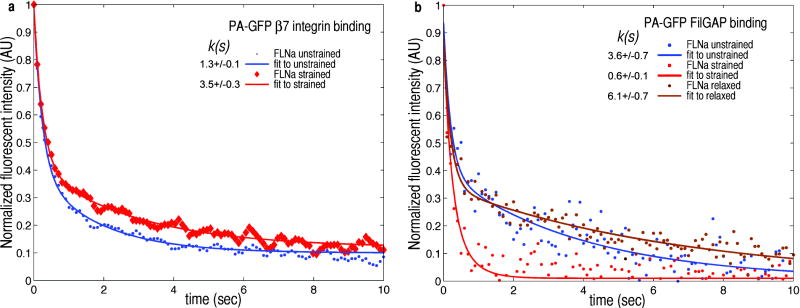
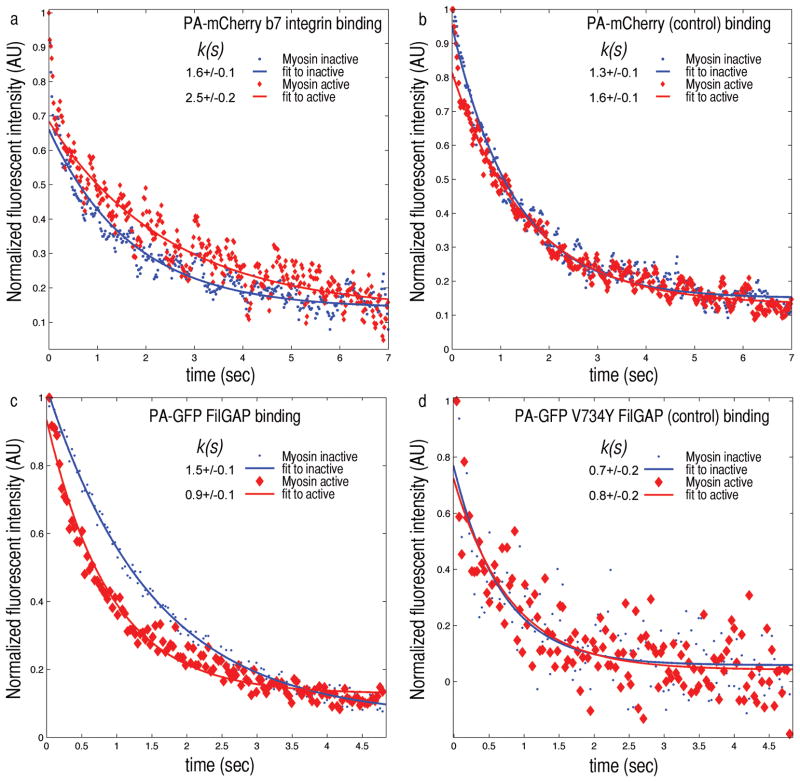
References
-
- Ingber DE. Mechanobiology and diseases of mechanotransduction. Ann Med. 2003;35:564–577. - PubMed
-
- Discher DE, Janmey P, Wang YL. Tissue cells feel and respond to the stiffness of their substrate. Science (New York, N Y. 2005;310:1139–1143. - PubMed
-
- Kruger M, Linke WA. Titin-based mechanical signalling in normal and failing myocardium. J Mol Cell Cardiol. 2009;46:490–498. S0022-2828(09)00021-2 [pii] - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
