

doi: 10.1007/7050_2007_026.
Spo11 and the Formation of DNA Double-Strand Breaks in Meiosis
Affiliations
- PMID: 21927624
- PMCID: PMC3172816
- DOI: 10.1007/7050_2007_026
Item in Clipboard
Spo11 and the Formation of DNA Double-Strand Breaks in Meiosis
Genome Dyn Stab.
.
Abstract
Meiotic recombination is carried out through a specialized pathway for the formation and repair of DNA double-strand breaks made by the Spo11 protein, a relative of archaeal topoisomerase VI. This review summarizes recent studies that provide insight to the mechanism of DNA cleavage by Spo11, functional interactions of Spo11 with other proteins required for break formation, mechanisms that control the timing of recombination initiation, and evolutionary conservation and divergence of these processes.
Figures
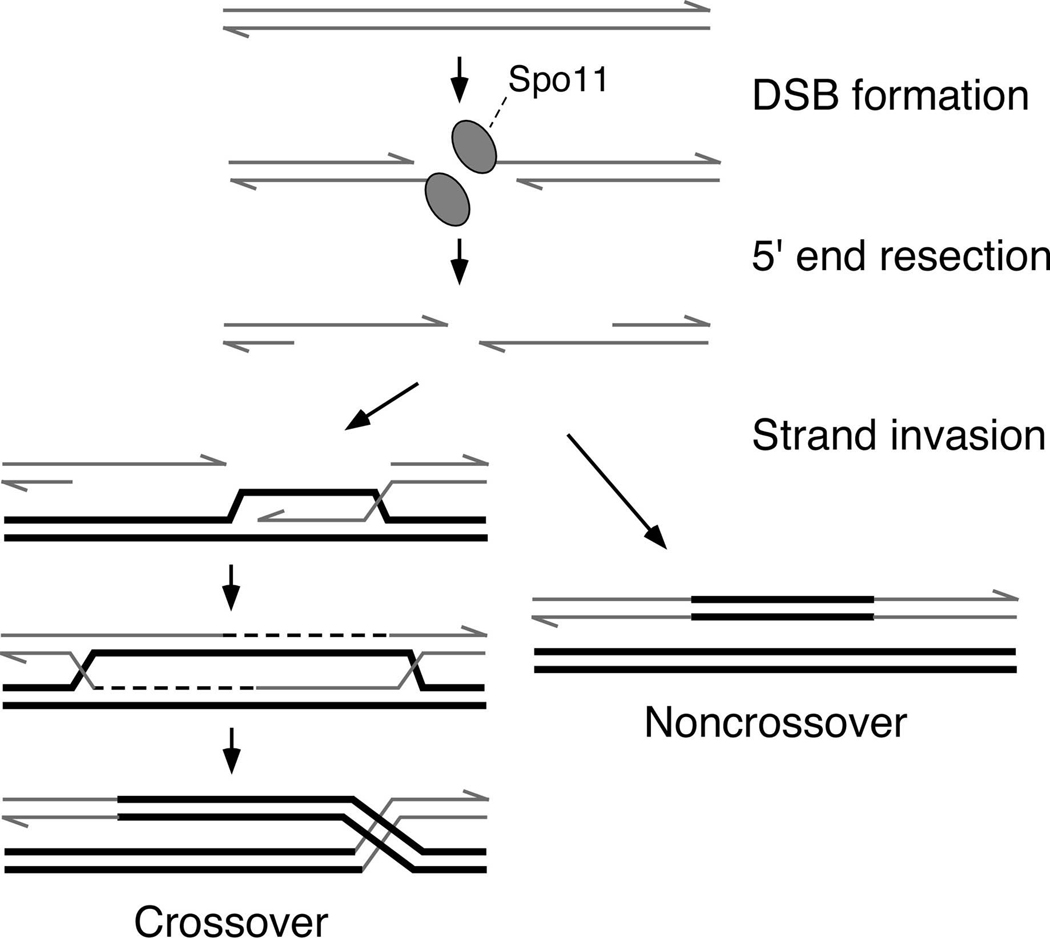
Schematic figures of the DNA at different steps (precursor, intermediates, and products) are shown. Proteins are omitted except for Spo11, which becomes covalently attached to the DNA as part of the DNA cleavage reaction. Only one of the two sister chromatids from each homolog. See text for details.
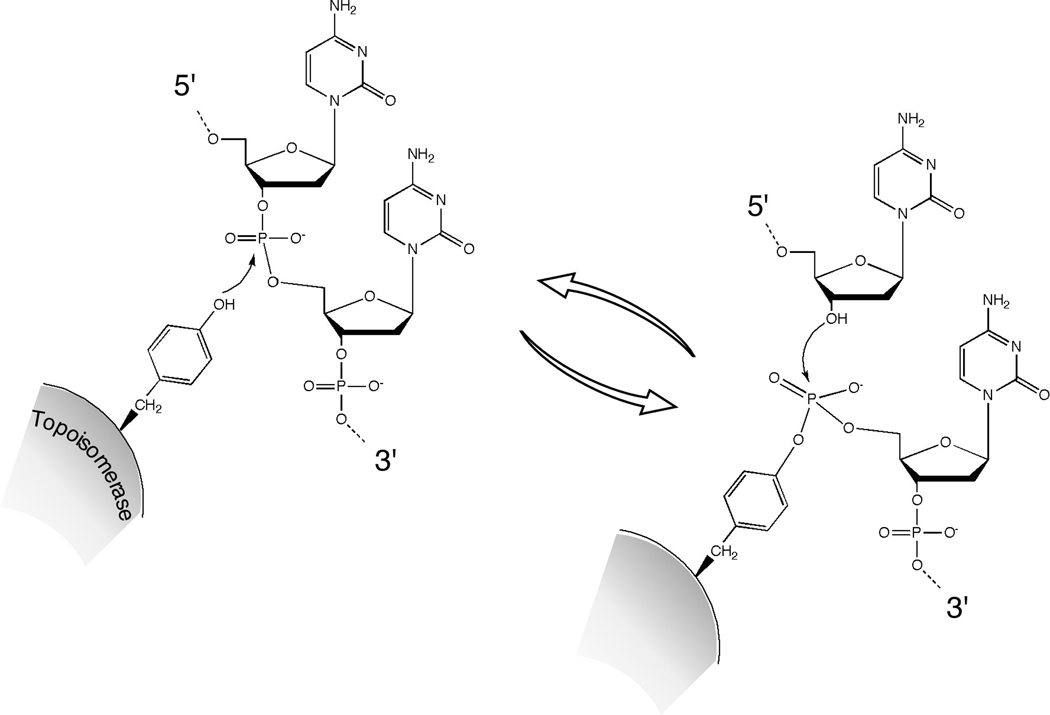
A tyrosine side chain on the topoisomerase protein carries out a nucleophilic attack on the DNA phosphodiester backbone. This transesterase reaction severs the DNA backbone and covalently links the protein to the DNA end via a tyrosyl phosphodiester linkage. The protein-DNA linkage is reversed and the DNA is resealed by attack of the deoxyribose hydroxyl. In this example, the tyrosyl phosphodiester links the protein to the 5′ end of the cleaved strand, as is seen in some type I and all type II topoisomerases. For a type II enzyme, two topoisomerase monomers work in concert to cleave the two strands of the duplex.
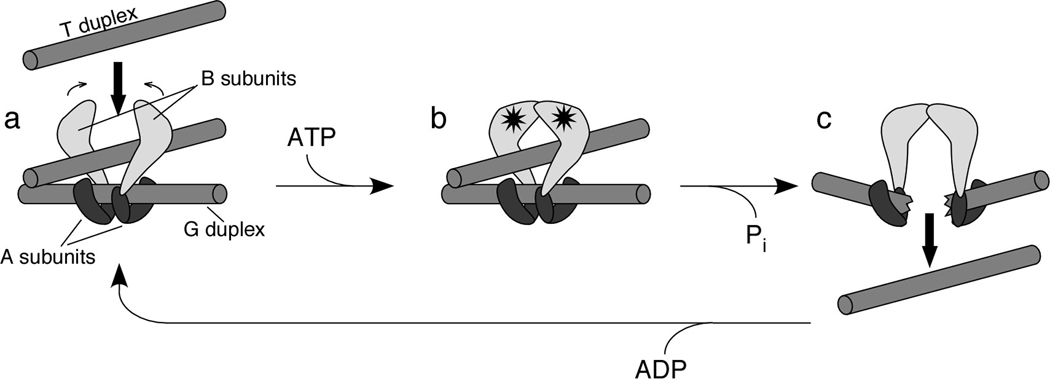
A two-gate model for TopoVI action has been proposed based on enzymatic and structural studies (Corbett and Berger 2004). Two B subunits are proposed to bind to a dimer of A subunits. The subunit-B binding interface on A is thought to reside on the CAP domain, which also contains the catalytic tyrosine residue. The dimer of A subunits binds to the DNA duplex that will be cleaved (the G, or gate, duplex). A second DNA duplex (the T, or transfer, duplex) enters the enzyme through a gate between the ATPase domains of the B subunits (panel a). ATP binding (stars) induces dimerization of the B subunits, trapping the T duplex (panel b). A conformational change accompanying ATP hydrolysis and DNA cleavage opens a gate in the G duplex, through which the T duplex is expelled (panel c). Release of ADP and resealing of the G duplex recycle the enzyme to its starting state. The figure is adapted, with permission, from the Annual Review of Biophysics and Biomolecular Structure, Vol. 33 (c)2004 by Annual Reviews (www.annualreviews.org ).
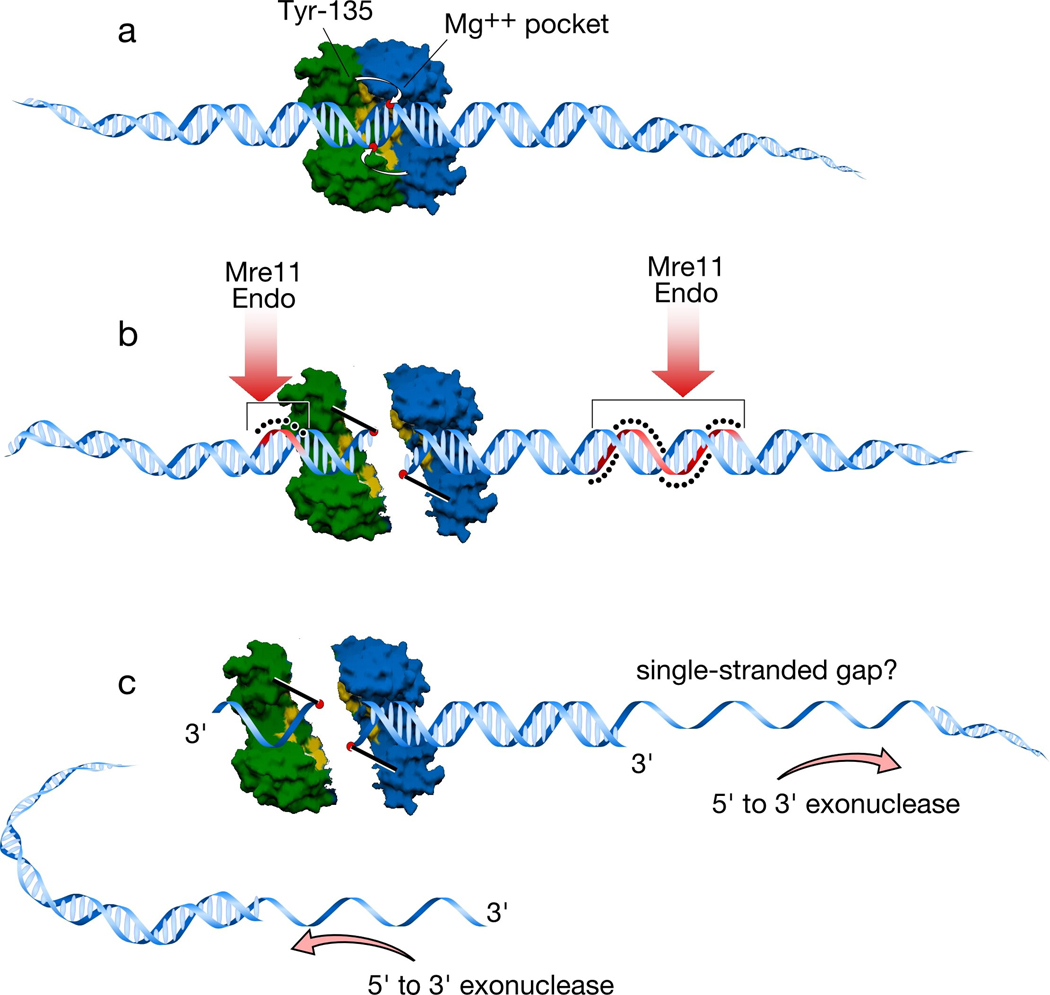
a. A Spo11 dimer is modeled using a surface rendering of the structure of Methanococcus jannaschi TopoVIA, with one monomer in green and the other in blue (Nichols et al. 1999). Approximate positions of the catalytic tyrosine of one monomer and the metal-binding pocket of the other monomer are indicated. B-form DNA (drawn approximately to scale) is docked onto the dimer. The metal binding pockets of the two monomers are appropriately spaced to place them in proximity to the scissile phosphates which would give the two-nucleotide 5′ overhang known to be generated by Spo11 and TopoVI. The tyrosines in the TopoVIA structure are far from these phosphates, however, so it is assumed that a conformational change in the protein moves the tyrosines into position to cleave (Nichols et al. 1999) (small white arrows). b. Cleavage of the DNA backbone leaves Spo11 covalently attached to the DSB ends. For clarity, the Spo11 dimer interface has been interrupted to show how the two monomers are attached. Analogous separation of the DNA ends is what opens a gate to allow strand passage for TopoVI (Corbett and Berger 2004); whether the Spo11 dimer interface might be disrupted in this manner is not known (see Section 5). After DNA cleavage, Spo11 is released from DSB ends by endonucleolytic single-strand cleavage on either side of the break, most likely mediated by Mre11 endonuclease activity. The strands where nicking occurs are colored red and highlighted with the black dots. Asymmetric nick spacing is shown, but the disposition of nicks around individual DSBs is not yet known. c. The DSB ends are further processed by 5′→3′ single-strand resection. The shorter Spo11-bound oligonucleotides may be small enough to be readily dissociated, but the longer oligonucleotides would need to be actively unwound. This feature may result in capping of one of the DSB ends, which may in turn influence the behavior of this end during subsequent recombination reactions (Neale et al. 2005).

The sequence conservation (% amino acid identity) between S. cerevisiae and each of several other Saccharomyces species is shown for the indicated proteins. DSB proteins diverge rapidly, especially when compared with two examples of mitotically expressed proteins, Rad51 and Arg4. Remarkably, Rec104 and Rec114 are less than 80% conserved even in S. paradoxus, a very close sibling of S. cerevisiae. A Rec104 homolog was not detected in the S. castellii genome sequence. Note that this analysis is not meant to imply that DSB proteins are uniquely prone to rapid divergence, although it has been demonstrated that meiotic recombination proteins in general tend to be among the cell's most divergent (Richard et al. 2005). Rather, the point is that they do diverge rapidly and may thus be difficult to identify by routine sequence homology searches in evolutionarily more distant organisms.

Spo11 is represented as for TopoVIA in Figure 3. The WD repeat protein Ski8 is shown as a donut bound to Spo11, Mre11 endonuclease as a pair of scissors, and other DSB proteins (“Rec102, etc.”) as clouds encompassing the Spo11 complex on DNA. Key features of this model are that Spo11 binding to DNA and activation to cleave are separate steps (Prieler et al. 2005); DNA cleavage by Spo11 and the onset of DSB processing by Mre11 (with or without other nucleases) are concerted reactions; and the irreversible Spo11-bound DSBs in rad50S and other mutants are off-pathway (Prieler et al. 2005). a. Assembly of a pre-DSB complex on DNA, consisting of Spo11, Ski8, and at least a subset of the other DSB proteins. b, c. Activation of Spo11 to cleave DNA. In panel b, conformational change within the complex (perhaps within Spo11 itself) renders Spo11 competent to cleave DNA, but it has not yet done so. This complex may be reversed without forming a DSB. In panel c, activated Spo11 complexes are competent to be driven forward: Spo11 forms a DSB and, in a concerted reaction, Mre11 nicks the DNA to initiate processing of the DSB. This sub-step is irreversible and commits this site to undergoing homologous recombination. d. Exonucleolytic processing. Further degradation of the 5′ terminal strands follows rapidly and in a concerted manner upon DSB formation and Mre11 nucleolytic activity. e. Irreversible DSB formation in rad50S, mre11S, and sae2 mutants. Activation of Spo11 in the absence of Mre11 nuclease activity stabilizes the complex (Prieler et al. 2005), but a frank DSB cannot form because concerted processing by Mre11 cannot occur. This complex eventually decays to give an irreversible DSB that is related to normal meiotic DSBs but that is off the normal pathway.
References
-
- Agashe B, Prasad CK, Siddiqi I. Identification and analysis of DYAD: a gene required for meiotic chromosome organisation and female meiotic progression in Arabidopsis. Development. 2002;129:3935–3943. - PubMed
-
- Alani E, Padmore R, Kleckner N. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell. 1990;61:419–436. - PubMed
-
- Arora C, Kee K, Maleki S, Keeney S. Antiviral protein Ski8 is a direct partner of Spo11 in meiotic DNA break formation, independent of its cytoplasmic role in RNA metabolism. Mol Cell. 2004;13:549–559. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources