

Adult neurogenesis in the decapod crustacean brain: a hematopoietic connection?
- PMID: 21929622
- PMCID: PMC3178839
- DOI: 10.1111/j.1460-9568.2011.07802.x
Adult neurogenesis in the decapod crustacean brain: a hematopoietic connection?
Abstract
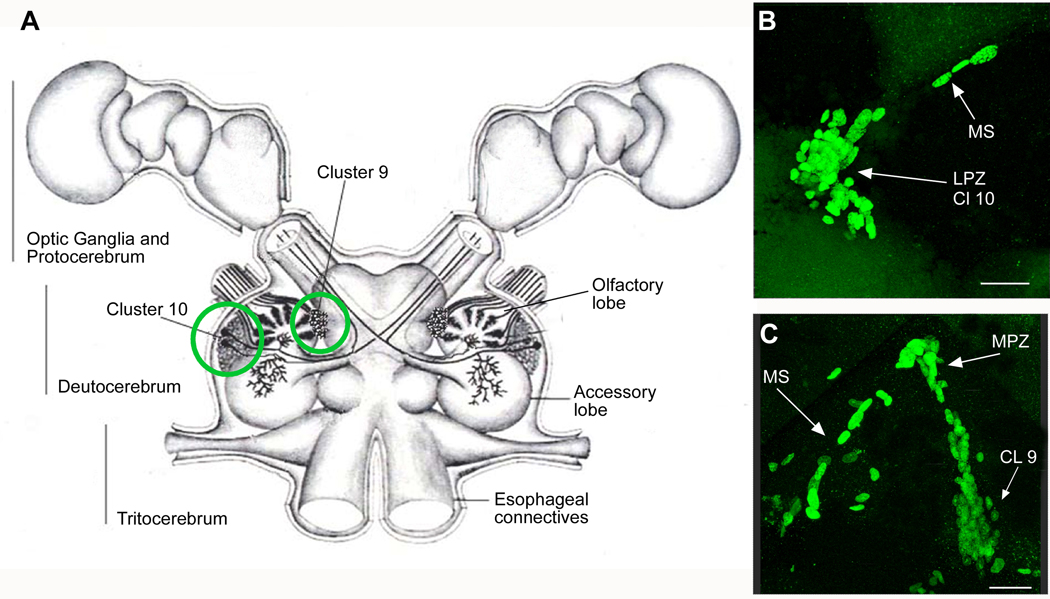
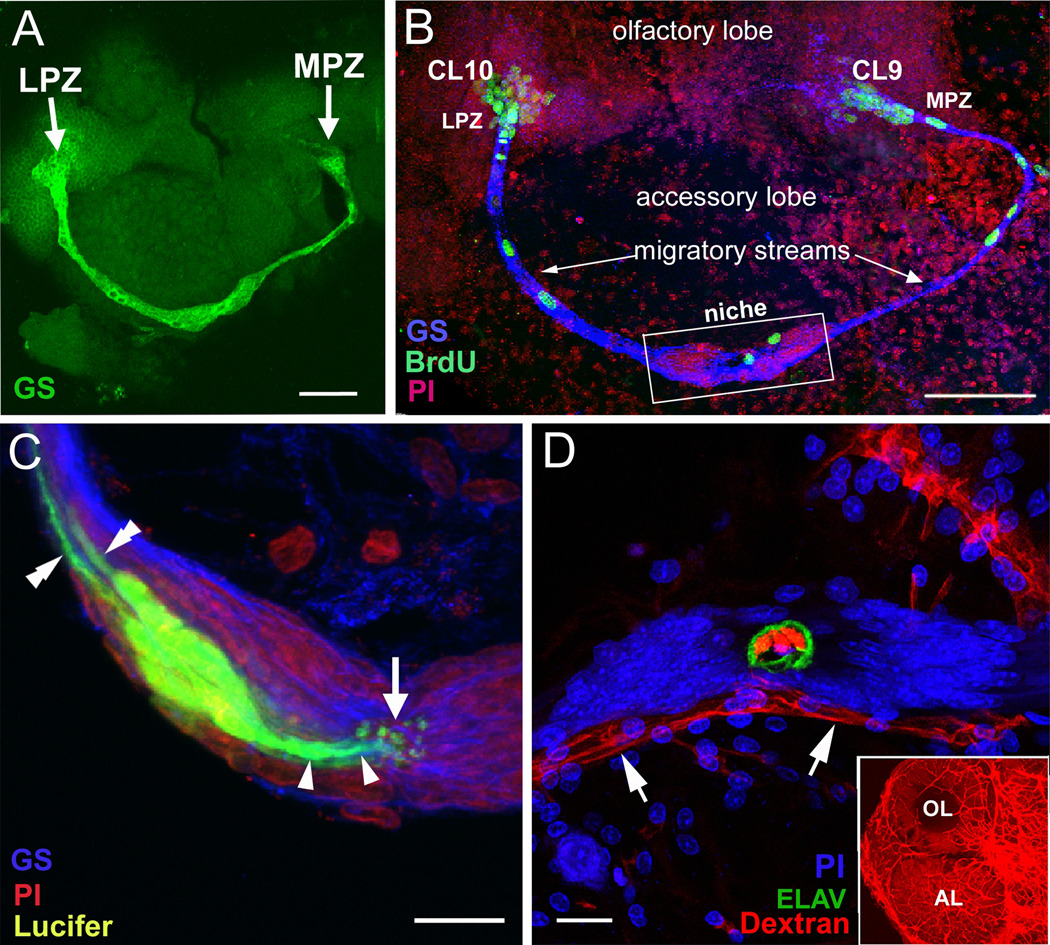
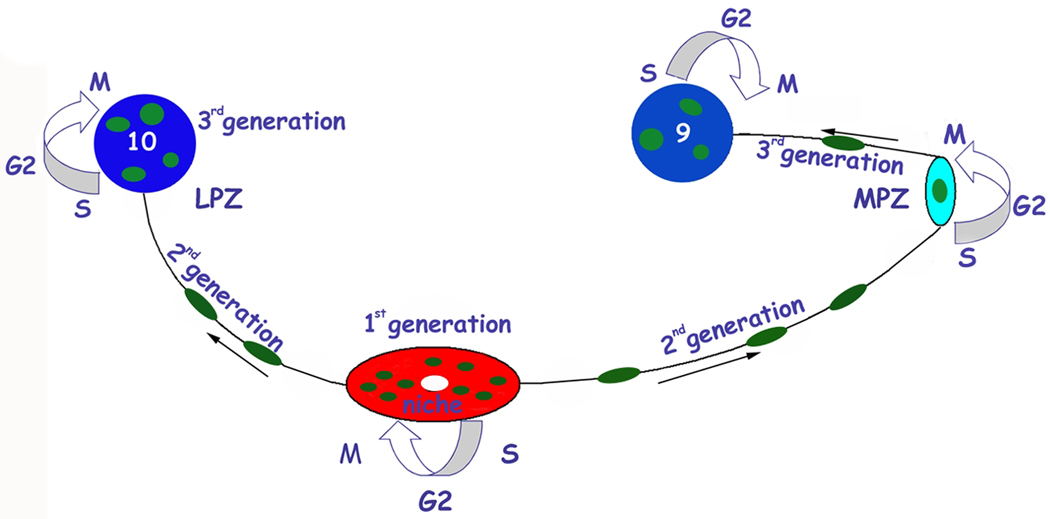
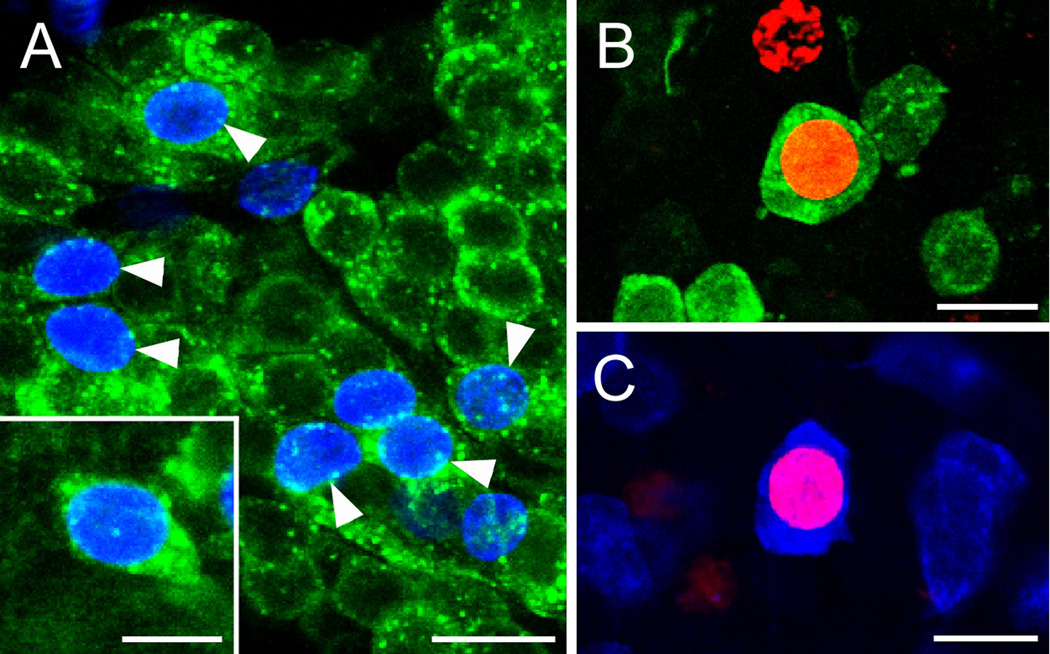
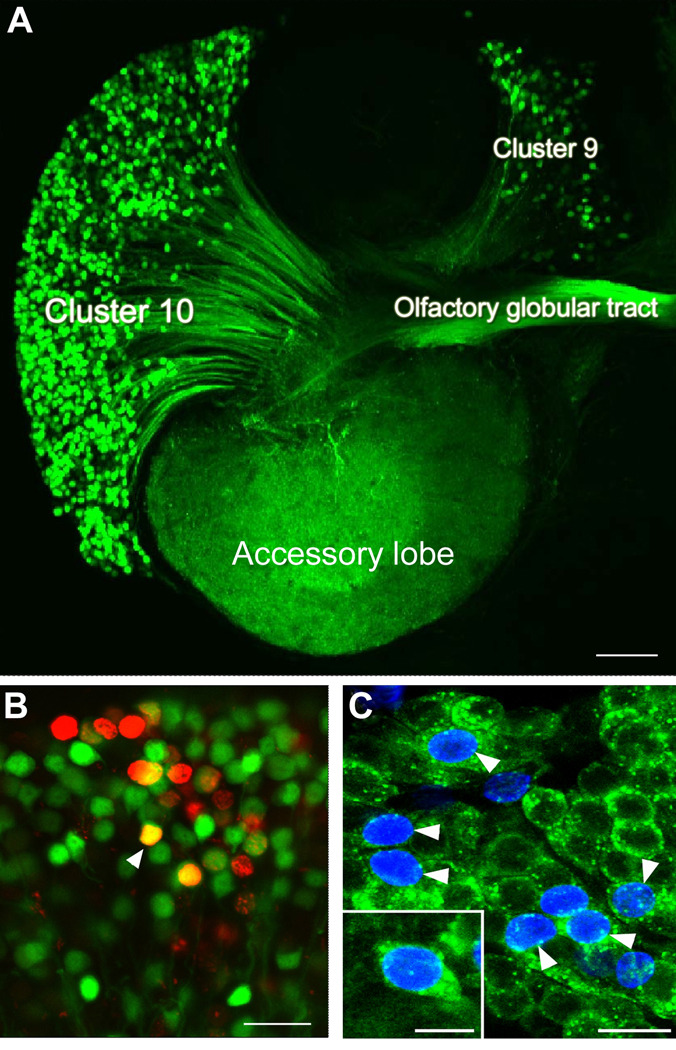
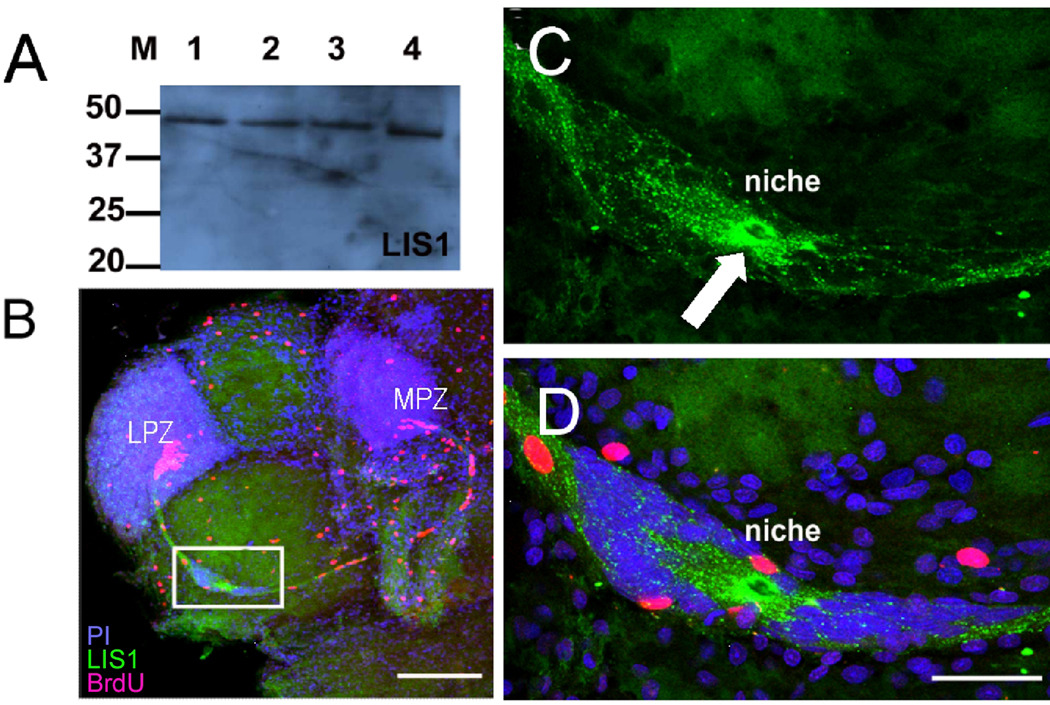
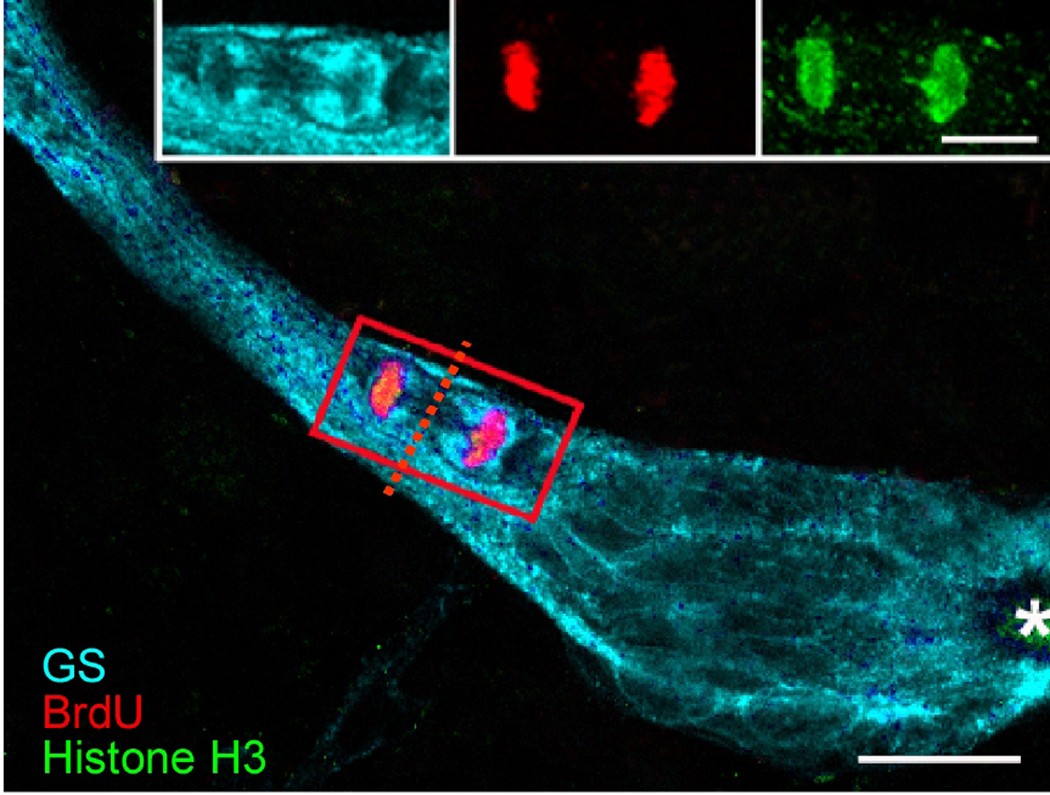
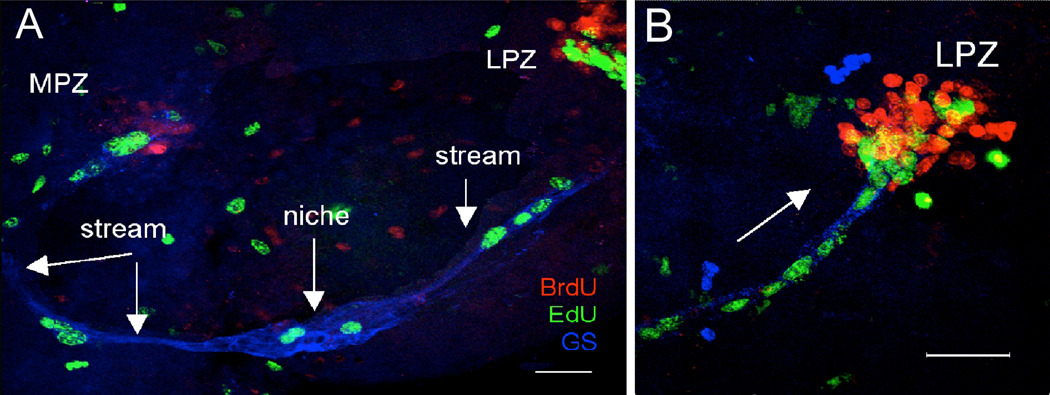
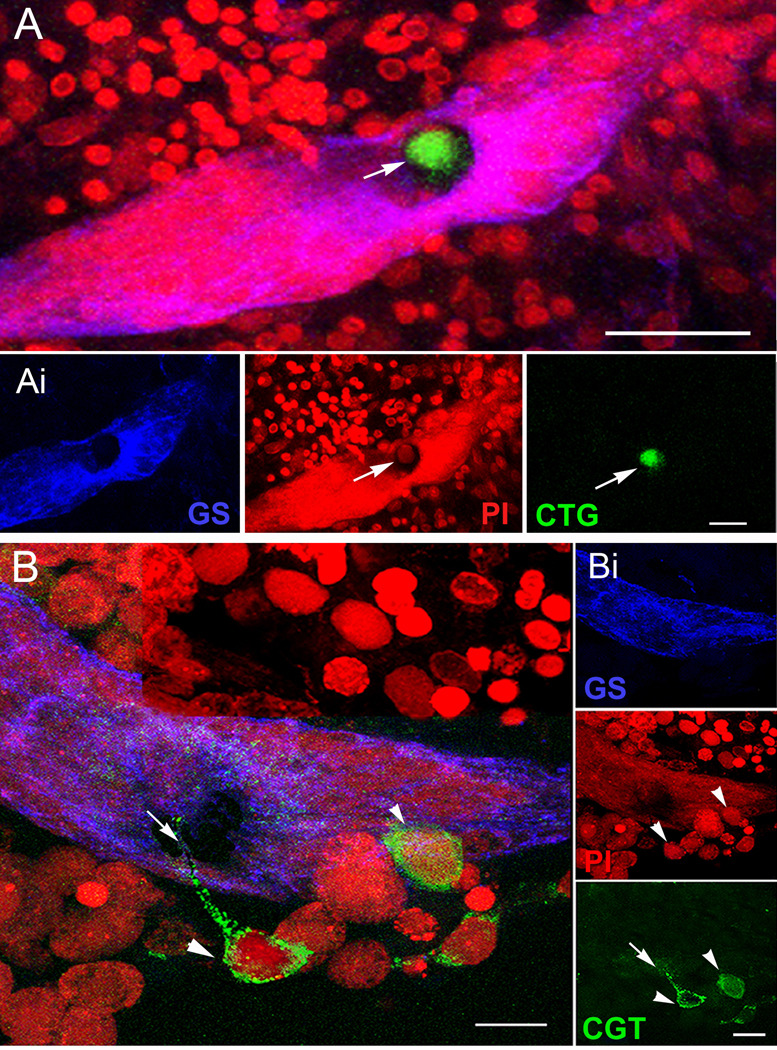
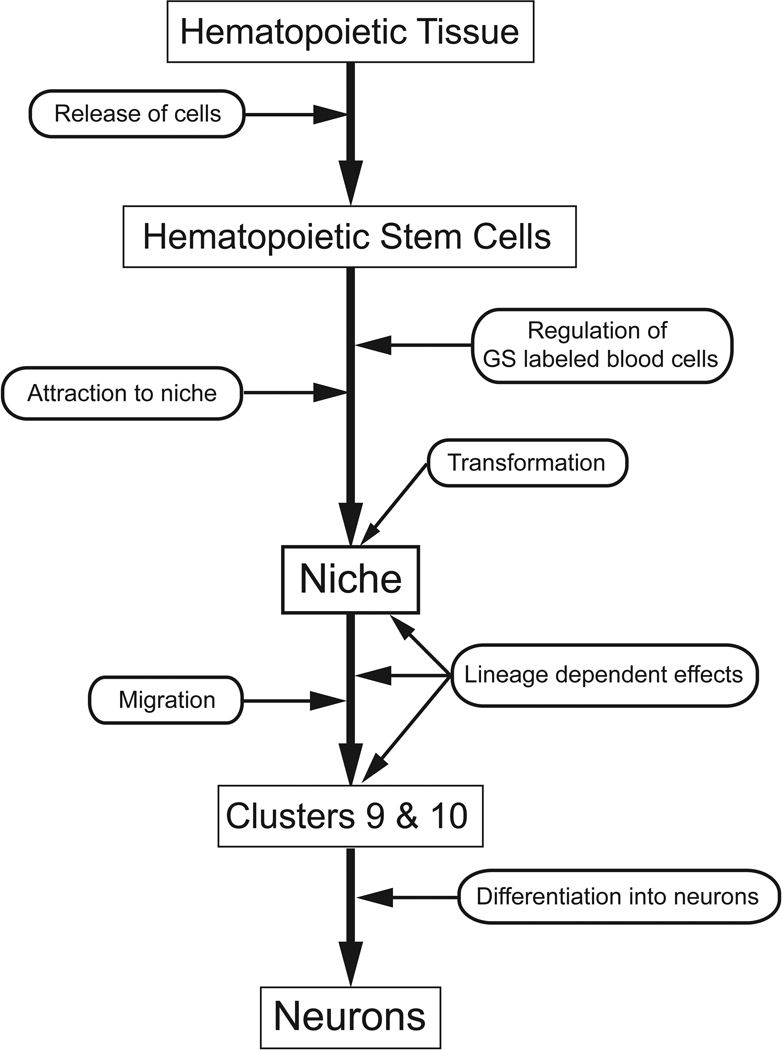
New neurons are produced and integrated into circuits in the adult brains of many organisms, including crustaceans. In some crustacean species, the first-generation neuronal precursors reside in a niche exhibiting characteristics analogous to mammalian neurogenic niches. However, unlike mammalian niches where several generations of neuronal precursors co-exist, the lineage of precursor cells in crayfish is spatially separated allowing the influence of environmental and endogenous regulators on specific generations in the neuronal precursor lineage to be defined. Experiments also demonstrate that the first-generation neuronal precursors in the crayfish Procambarus clarkii are not self-renewing. A source external to the neurogenic niche must therefore provide cells that replenish the first-generation precursor pool, because although these cells divide and produce a continuous efflux of second-generation cells from the niche, the population of first-generation niche precursors is not diminished with growth and aging. In vitro studies show that cells extracted from the hemolymph, but not other tissues, are attracted to and incorporated into the neurogenic niche, a phenomenon that appears to involve serotonergic mechanisms. We propose that, in crayfish, the hematopoietic system may be a source of cells that replenish the niche cell pool. These and other studies reviewed here establish decapod crustaceans as model systems in which the processes underlying adult neurogenesis, such as stem cell origins and transformation, can be readily explored. Studies in diverse species where adult neurogenesis occurs will result in a broader understanding of fundamental mechanisms and how evolutionary processes may have shaped the vertebrate/mammalian condition.
© 2011 The Authors. European Journal of Neuroscience © 2011 Federation of European Neuroscience Societies and Blackwell Publishing Ltd.
Figures
References
-
- Bazin F. Étude comparée de l’organe deutocérébral des Macroures Reptantia et des Anomoures (Crustacés Décapodes) Arch. Zool. Exp. Gen. 1970a;111:245–264.
-
- Bazin F. Les organs deutocérébraux chez deux Crustacés Décapodes Macroures Reptantia: Panulirus regius de Brito Capello, Scyllarus arctus (L.) B. Soc. Zool. Fr. 1970b;96:87–92.
-
- Bazin F, Demeusy N. Existance d’organes intracérébraux énigmatiques chez le Crustacé Décapode Carcinus maenas (L.) C. R. Acad. Sci. 1968;267:356–358.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
