

Protein phosphatase 2A (PP2A)-specific ubiquitin ligase MID1 is a sequence-dependent regulator of translation efficiency controlling 3-phosphoinositide-dependent protein kinase-1 (PDPK-1)
- PMID: 21930711
- PMCID: PMC3220588
- DOI: 10.1074/jbc.M111.224451
Protein phosphatase 2A (PP2A)-specific ubiquitin ligase MID1 is a sequence-dependent regulator of translation efficiency controlling 3-phosphoinositide-dependent protein kinase-1 (PDPK-1)
Abstract
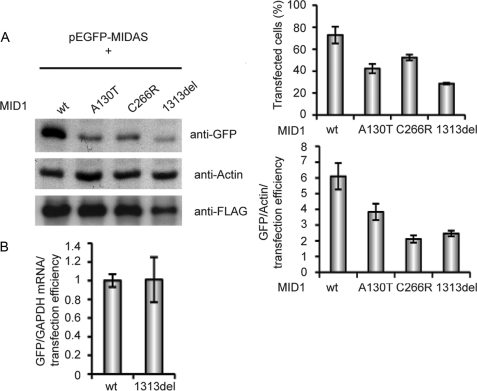
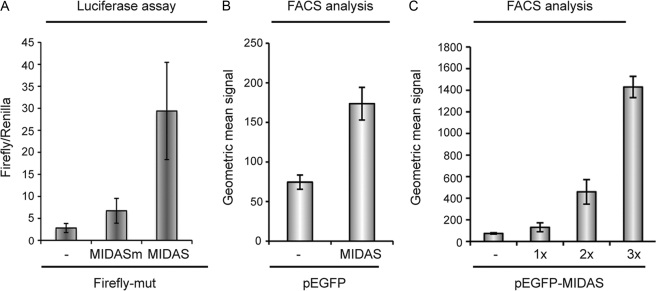
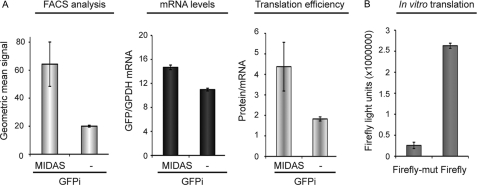
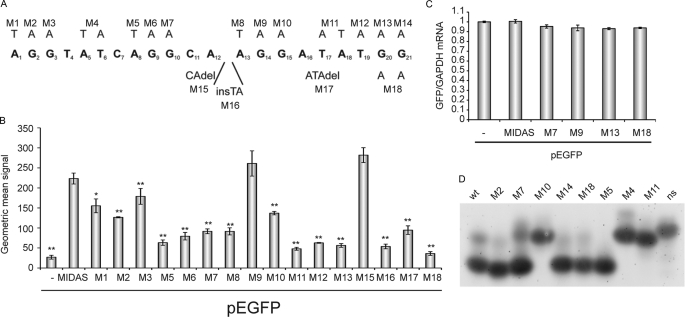
We have shown previously that the ubiquitin ligase MID1, mutations of which cause the midline malformation Opitz BBB/G syndrome (OS), serves as scaffold for a microtubule-associated protein complex that regulates protein phosphatase 2A (PP2A) activity in a ubiquitin-dependent manner. Here, we show that the MID1 protein complex associates with mRNAs via a purine-rich sequence motif called MIDAS (MID1 association sequence) and thereby increases stability and translational efficiency of these mRNAs. Strikingly, inclusion of multiple copies of the MIDAS motif into mammalian mRNAs increases production of the encoded proteins up to 20-fold. Mutated MID1, as found in OS patients, loses its influence on MIDAS-containing mRNAs, suggesting that the malformations in OS patients could be caused by failures in the regulation of cytoskeleton-bound protein translation. This is supported by the observation that the majority of mRNAs that carry MIDAS motifs is involved in developmental processes and/or energy homeostasis. Further analysis of one of the proteins encoded by a MIDAS-containing mRNA, namely PDPK-1 (3-phosphoinositide dependent protein kinase-1), which is an important regulator of mammalian target of rapamycin/PP2A signaling, showed that PDPK-1 protein synthesis is significantly reduced in cells from an OS patient compared with an age-matched control and can be rescued by functional MID1. Together, our data uncover a novel messenger ribonucleoprotein complex that regulates microtubule-associated protein translation. They suggest a novel mechanism underlying OS and point at an enormous potential of the MIDAS motif to increase the efficiency of biotechnological protein production in mammalian cells.
Figures



References
-
- Quaderi N. A., Schweiger S., Gaudenz K., Franco B., Rugarli E. I., Berger W., Feldman G. J., Volta M., Andolfi G., Gilgenkrantz S., Marion R. W., Hennekam R. C., Opitz J. M., Muenke M., Ropers H. H., Ballabio A. (1997) Nat. Genet. 17, 285–291 - PubMed
-
- Inui S., Sanjo H., Maeda K., Yamamoto H., Miyamoto E., Sakaguchi N. (1998) Blood 92, 539–546 - PubMed
-
- Trockenbacher A., Suckow V., Foerster J., Winter J., Krauss S., Ropers H. H., Schneider R., Schweiger S. (2001) Nat. Genet. 29, 287–294 - PubMed
-
- Jacinto E., Hall M. N. (2003) Nat. Rev. Mol. Cell Biol. 4, 117–126 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
