

Candida albicans cell wall glycosylation may be indirectly required for activation of epithelial cell proinflammatory responses
- PMID: 21930756
- PMCID: PMC3232641
- DOI: 10.1128/IAI.05591-11
Candida albicans cell wall glycosylation may be indirectly required for activation of epithelial cell proinflammatory responses
Abstract
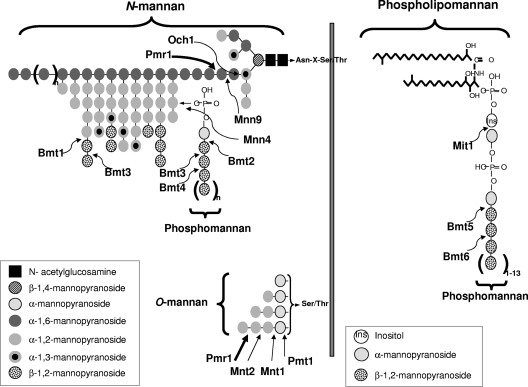
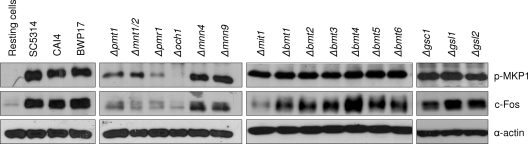
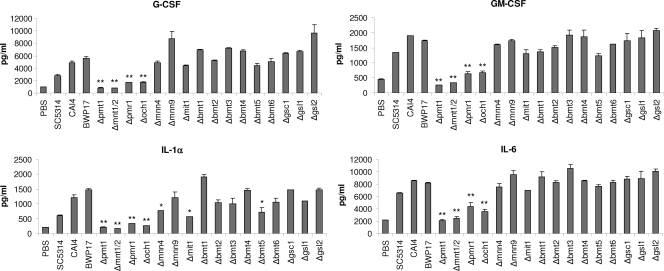
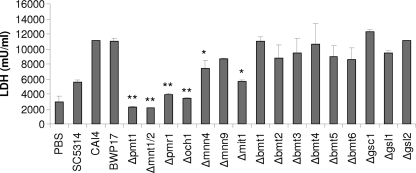
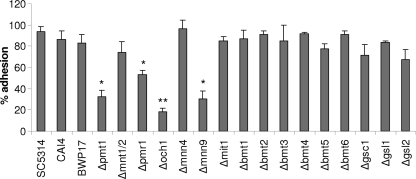
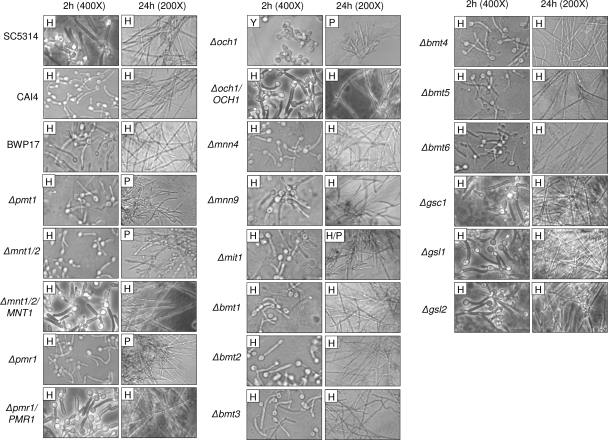
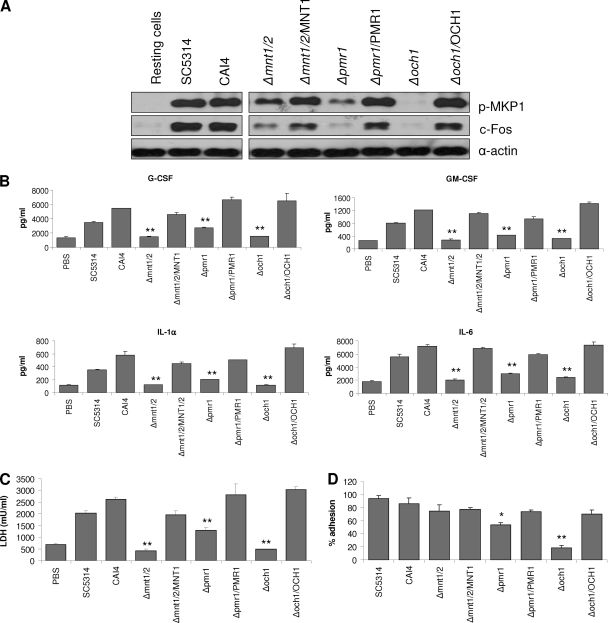
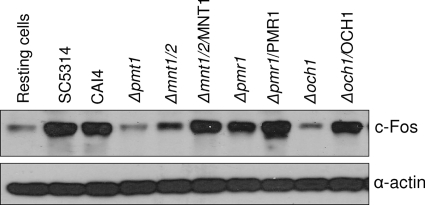
Oral epithelial cells discriminate between the yeast and hyphal forms of Candida albicans via the mitogen-activated protein kinase (MAPK) signaling pathway. This occurs through phosphorylation of the MAPK phosphatase MKP1 and activation of the c-Fos transcription factor by the hyphal form. Given that fungal cell wall polysaccharides are critical in host recognition and immune activation in myeloid cells, we sought to determine whether β-glucan and N- or O-glycosylation was important in activating the MAPK/MKP1/c-Fos hypha-mediated response mechanism and proinflammatory cytokines in oral epithelial cells. Using a series of β-glucan and N- and O-mannan mutants, we found that N-mannosylation (via Δoch1 and Δpmr1 mutants) and O-mannosylation (via Δpmt1 and Δmnt1 Δmnt2 mutants), but not phosphomannan (via a Δmnn4 mutant) or β-1,2 mannosylation (via Δbmt1 to Δbmt6 mutants), were required for MKP1/c-Fos activation, proinflammatory cytokine production, and cell damage induction. However, the N- and O-mannan mutants showed reduced adhesion or lack of initial hypha formation at 2 h, resulting in little MKP1/c-Fos activation, or restricted hypha formation/pseudohyphal formation at 24 h, resulting in minimal proinflammatory cytokine production and cell damage. Further, the α-1,6-mannose backbone of the N-linked outer chain (corresponding to a Δmnn9 mutant) may be required for epithelial adhesion, while the α-1,2-mannose component of phospholipomannan (corresponding to a Δmit1 mutant) may contribute to epithelial cell damage. β-Glucan appeared to play no role in adhesion, epithelial activation, or cell damage. In summary, N- and O-mannosylation defects affect the ability of C. albicans to induce proinflammatory cytokines and damage in oral epithelial cells, but this may be due to indirect effects on fungal pathogenicity rather than mannose residues being direct activators of the MAPK/MKP1/c-Fos hypha-mediated immune response.
Figures
References
-
- Bates S., et al. 2005. Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J. Biol. Chem. 280:23408–23415 - PubMed
-
- Bates S., et al. 2006. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J. Biol. Chem. 281:90–98 - PubMed
-
- Brown G. D. 2006. Dectin-1: a signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 6:33–43 - PubMed
-
- Cambi A., et al. 2003. The C-type lectin DC-SIGN (CD209) is an antigen-uptake receptor for Candida albicans on dendritic cells. Eur. J. Immunol. 33:532–538 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
