

A genomic model of condition-specific nucleosome behavior explains transcriptional activity in yeast
- PMID: 21930892
- PMCID: PMC3246209
- DOI: 10.1101/gr.124099.111
A genomic model of condition-specific nucleosome behavior explains transcriptional activity in yeast
Abstract
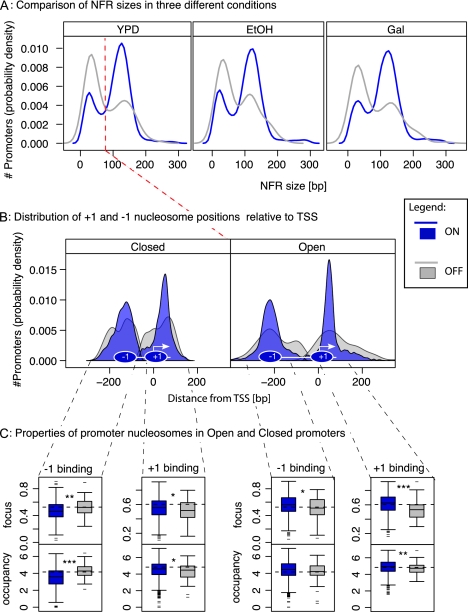
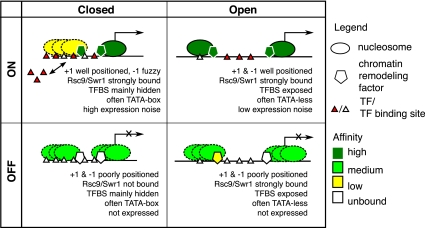
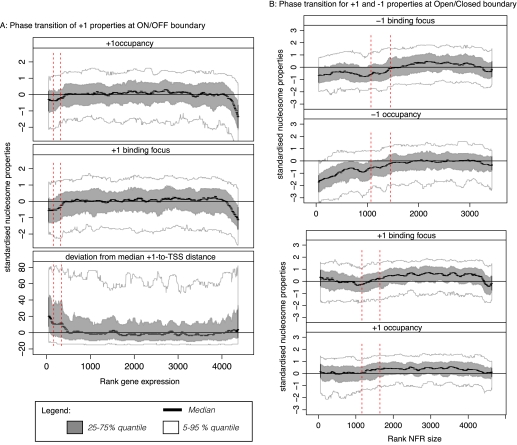
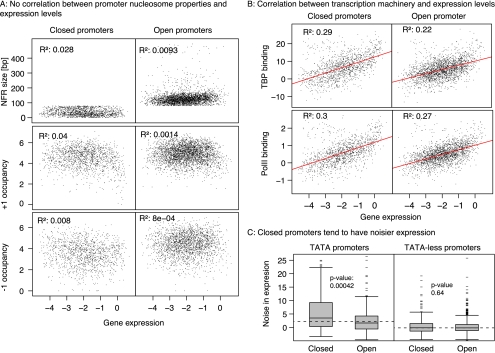
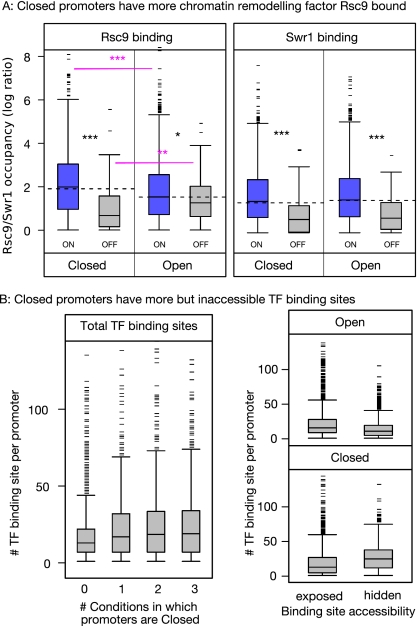
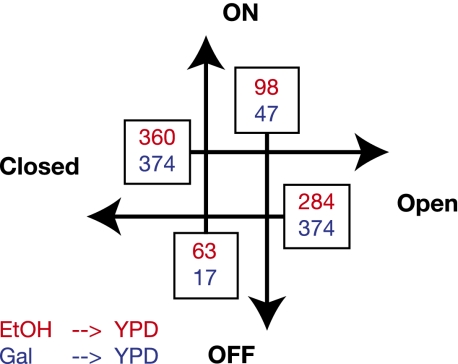
Nucleosomes play an important role in gene regulation. Molecular studies observed that nucleosome binding in promoters tends to be repressive. In contrast, genomic studies have delivered conflicting results: An analysis of yeast grown on diverse carbon sources reported that nucleosome occupancies remain largely unchanged between conditions, whereas a study of the heat-shock response suggested that nucleosomes get evicted at promoters of genes with increased expression. Consequently, there are few general principles that capture the relationship between chromatin organization and transcriptional regulation. Here, we present a qualitative model for nucleosome positioning in Saccharomyces cerevisiae that helps explain important properties of gene expression. By integrating publicly available data sets, we observe that promoter-bound nucleosomes assume one of four discrete configurations that determine the active and silent transcriptional states of a gene, but not its expression level. In TATA-box-containing promoters, nucleosome architecture indicates the amount of transcriptional noise. We show that >20% of genes switch promoter states upon changes in cellular conditions. The data suggest that DNA-binding transcription factors together with chromatin-remodeling enzymes are primarily responsible for the nucleosome architecture. Our model for promoter nucleosome architecture reconciles genome-scale findings with molecular studies; in doing so, we establish principles for nucleosome positioning and gene expression that apply not only to individual genes, but across the entire genome. The study provides a stepping stone for future models of transcriptional regulation that encompass the intricate interplay between cis- and trans-acting factors, chromatin, and the core transcriptional machinery.
Figures
References
-
- Adkins MW, Tyler JK 2006. Transcriptional activators are dispensable for transcription in the absence of Spt6-mediated chromatin reassembly of promoter regions. Mol Cell 21: 405–416 - PubMed
-
- Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, Schuster SC, Pugh BF 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446: 572–576 - PubMed
-
- Basehoar AD, Zanton SJ, Pugh BF 2004. Identification and distinct regulation of yeast TATA box-containing genes. Cell 116: 699–709 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases