

Nongenomic glucocorticoid receptor action regulates gap junction intercellular communication and neural progenitor cell proliferation
- PMID: 21930911
- PMCID: PMC3189065
- DOI: 10.1073/pnas.1102821108
Nongenomic glucocorticoid receptor action regulates gap junction intercellular communication and neural progenitor cell proliferation
Abstract
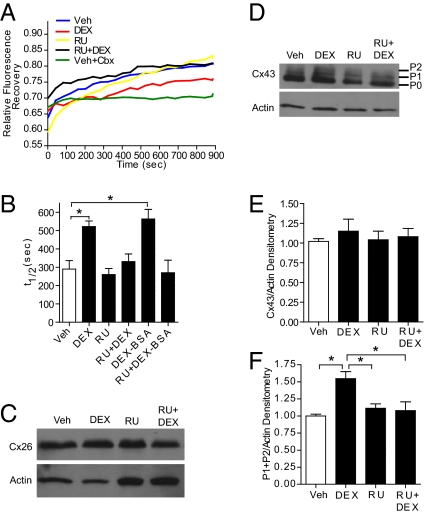
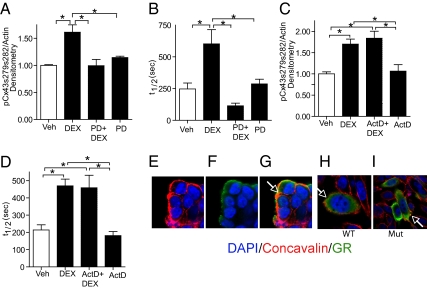
Glucocorticoids (GCs) are used to treat pregnant women at risk for preterm delivery; however, prenatal exposure to GCs may trigger adverse neurological side effects due to reduced neural progenitor cell (NPC) proliferation. Whereas many established cell-cycle regulators impact NPC proliferation, other signaling molecules, such as the gap junction protein connexin-43 (Cx43), also influence proliferation. Gap junction intercellular communication (GJIC) is influenced by GCs in some cells, but such hormone effects have not been examined in coupled stem cells. We found that both continuous and transient exposure of embryonic day 14.5 mouse neurosphere cultures to dexamethasone (DEX) limits proliferation of coupled NPCs, which is manifested by both a reduction in S-phase progression and enhanced cell-cycle exit. A short (i.e., 1-h) DEX treatment also reduced GJIC as measured by live-cell fluorescence recovery after photobleaching, and altered the synchrony of spontaneous calcium transients in coupled NPCs. GC effects on GJIC in NPCs are transcription-independent and mediated through plasma membrane glucocorticoid receptors (GRs). This nongenomic pathway operates through lipid raft-associated GRs via a site-specific, MAPK-dependent phosphorylation of Cx43, which is linked to GR via caveolin-1 (Cav-1) and c-src. Cav-1 is essential for this nongenomic action of GR, as DEX effects on GJIC, Cx43 phosphorylation, and MAPK activation are not observed in Cav-1 knockout NPCs. As transient pharmacologic inhibition of GJIC triggers reduced S-phase progression but not enhanced cell-cycle exit, the nongenomic GR signaling pathway may operate via distinct downstream effectors to alter the proliferative capacity of NPCs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Caveolin-1 regulates genomic action of the glucocorticoid receptor in neural stem cells.Mol Cell Biol. 2014 Jul;34(14):2611-23. doi: 10.1128/MCB.01121-13. Mol Cell Biol. 2014. PMID: 24777604 Free PMC article.
-
Involvement of connexin43 in the EGF/EGFR signalling during self-renewal and differentiation of neural progenitor cells.Cell Signal. 2013 Dec;25(12):2676-84. doi: 10.1016/j.cellsig.2013.08.030. Epub 2013 Sep 4. Cell Signal. 2013. PMID: 24012498
-
Glucocorticoid Suppresses Connexin 43 Expression by Inhibiting the Akt/mTOR Signaling Pathway in Osteoblasts.Calcif Tissue Int. 2016 Jul;99(1):88-97. doi: 10.1007/s00223-016-0121-y. Epub 2016 Feb 25. Calcif Tissue Int. 2016. PMID: 26914606
-
Cooperativity and complementarity: synergies in non-classical and classical glucocorticoid signaling.Cell Cycle. 2012 Aug 1;11(15):2819-27. doi: 10.4161/cc.21018. Epub 2012 Aug 1. Cell Cycle. 2012. PMID: 22801547 Free PMC article. Review.
-
Mind the gap; regulation of gap junctional, intercellular communication by the SRC oncogene product and its effectors.Anticancer Res. 2012 Oct;32(10):4245-50. Anticancer Res. 2012. PMID: 23060544 Review.
Cited by
-
Topical Dexamethasone Administration Impairs Protein Synthesis and Neuronal Regeneration in the Olfactory Epithelium.Front Mol Neurosci. 2018 Mar 6;11:50. doi: 10.3389/fnmol.2018.00050. eCollection 2018. Front Mol Neurosci. 2018. PMID: 29559887 Free PMC article.
-
The biology of the glucocorticoid receptor: new signaling mechanisms in health and disease.J Allergy Clin Immunol. 2013 Nov;132(5):1033-44. doi: 10.1016/j.jaci.2013.09.007. Epub 2013 Sep 29. J Allergy Clin Immunol. 2013. PMID: 24084075 Free PMC article. Review.
-
Corticosteroids: Mechanisms of Action in Health and Disease.Rheum Dis Clin North Am. 2016 Feb;42(1):15-31, vii. doi: 10.1016/j.rdc.2015.08.002. Rheum Dis Clin North Am. 2016. PMID: 26611548 Free PMC article. Review.
-
Genomic glucocorticoid action in embryonic mouse neural stem cells.Mol Cell Endocrinol. 2023 Mar 1;563:111864. doi: 10.1016/j.mce.2023.111864. Epub 2023 Jan 20. Mol Cell Endocrinol. 2023. PMID: 36690169 Free PMC article.
-
Statins impact primary embryonic mouse neural stem cell survival, cell death, and fate through distinct mechanisms.PLoS One. 2018 May 8;13(5):e0196387. doi: 10.1371/journal.pone.0196387. eCollection 2018. PLoS One. 2018. PMID: 29738536 Free PMC article.
References
-
- Haller J, Mikics E, Makara GB. The effects of non-genomic glucocorticoid mechanisms on bodily functions and the central neural system. A critical evaluation of findings. Front Neuroendocrinol. 2008;29:273–291. - PubMed
-
- Pasricha N, Joëls M, Karst H. Rapid effects of corticosterone in the mouse dentate gyrus via a nongenomic pathway. J Neuroendocrinol. 2011;23:143–147. - PubMed
-
- Qiu J, et al. Rapid activation of ERK1/2 mitogen-activated protein kinase by corticosterone in PC12 cells. Biochem Biophys Res Commun. 2001;287:1017–1024. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
