

α-Galactosidase/sucrose kinase (AgaSK), a novel bifunctional enzyme from the human microbiome coupling galactosidase and kinase activities
- PMID: 21931163
- PMCID: PMC3220449
- DOI: 10.1074/jbc.M111.286039
α-Galactosidase/sucrose kinase (AgaSK), a novel bifunctional enzyme from the human microbiome coupling galactosidase and kinase activities
Abstract
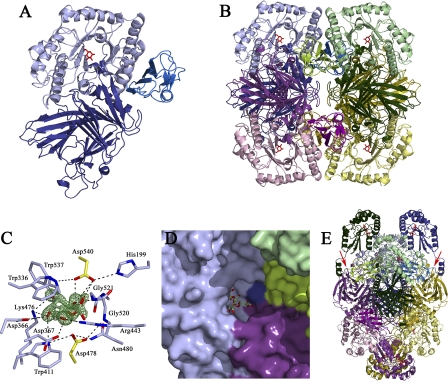
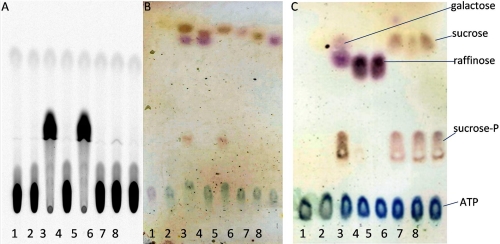
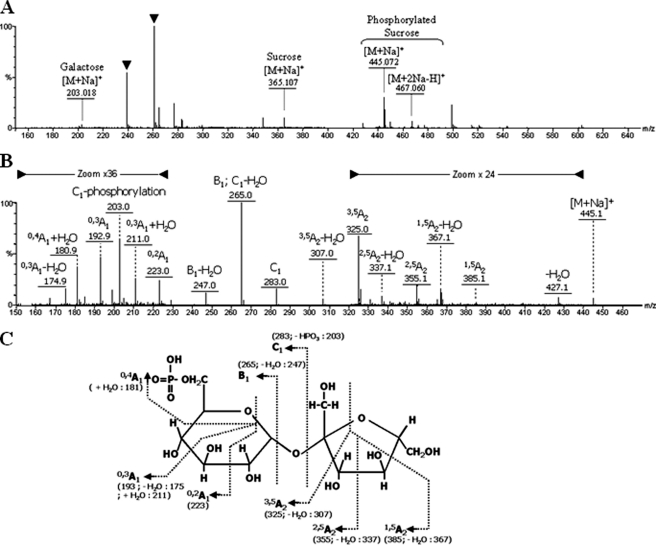
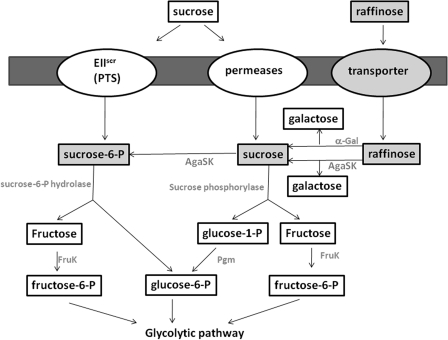
α-Galactosides are non-digestible carbohydrates widely distributed in plants. They are a potential source of energy in our daily food, and their assimilation by microbiota may play a role in obesity. In the intestinal tract, they are degraded by microbial glycosidases, which are often modular enzymes with catalytic domains linked to carbohydrate-binding modules. Here we introduce a bifunctional enzyme from the human intestinal bacterium Ruminococcus gnavus E1, α-galactosidase/sucrose kinase (AgaSK). Sequence analysis showed that AgaSK is composed of two domains: one closely related to α-galactosidases from glycoside hydrolase family GH36 and the other containing a nucleotide-binding motif. Its biochemical characterization showed that AgaSK is able to hydrolyze melibiose and raffinose to galactose and either glucose or sucrose, respectively, and to specifically phosphorylate sucrose on the C6 position of glucose in the presence of ATP. The production of sucrose-6-P directly from raffinose points toward a glycolytic pathway in bacteria, not described so far. The crystal structures of the galactosidase domain in the apo form and in complex with the product shed light onto the reaction and substrate recognition mechanisms and highlight an oligomeric state necessary for efficient substrate binding and suggesting a cross-talk between the galactose and kinase domains.
Figures
Similar articles
-
α-Galactosidase and Sucrose-Kinase Relationships in a Bi-functional AgaSK Enzyme Produced by the Human Gut Symbiont Ruminococcus gnavus E1.Front Microbiol. 2020 Nov 12;11:579521. doi: 10.3389/fmicb.2020.579521. eCollection 2020. Front Microbiol. 2020. PMID: 33281771 Free PMC article.
-
Functional analysis of family GH36 α-galactosidases from Ruminococcus gnavus E1: insights into the metabolism of a plant oligosaccharide by a human gut symbiont.Appl Environ Microbiol. 2012 Nov;78(21):7720-32. doi: 10.1128/AEM.01350-12. Epub 2012 Aug 24. Appl Environ Microbiol. 2012. PMID: 22923411 Free PMC article.
-
Crystal structure of α-galactosidase from Lactobacillus acidophilus NCFM: insight into tetramer formation and substrate binding.J Mol Biol. 2011 Sep 23;412(3):466-80. doi: 10.1016/j.jmb.2011.07.057. Epub 2011 Jul 30. J Mol Biol. 2011. PMID: 21827767
-
The melREDCA Operon Encodes a Utilization System for the Raffinose Family of Oligosaccharides in Bacillus subtilis.J Bacteriol. 2019 Jul 10;201(15):e00109-19. doi: 10.1128/JB.00109-19. Print 2019 Aug 1. J Bacteriol. 2019. PMID: 31138628 Free PMC article.
-
Oligomeric Structure of Yeast and Other Invertases Governs Specificity.Subcell Biochem. 2024;104:503-530. doi: 10.1007/978-3-031-58843-3_19. Subcell Biochem. 2024. PMID: 38963498 Review.
Cited by
-
Activity-based metaproteomics driven discovery and enzymological characterization of potential α-galactosidases in the mouse gut microbiome.Commun Chem. 2024 Aug 16;7(1):184. doi: 10.1038/s42004-024-01273-5. Commun Chem. 2024. PMID: 39152233 Free PMC article.
-
Structural elucidation of dextran degradation mechanism by streptococcus mutans dextranase belonging to glycoside hydrolase family 66.J Biol Chem. 2012 Jun 8;287(24):19916-26. doi: 10.1074/jbc.M112.342444. Epub 2012 Feb 15. J Biol Chem. 2012. PMID: 22337884 Free PMC article.
-
RadA, a MSCRAMM Adhesin of the Dominant Symbiote Ruminococcus gnavus E1, Binds Human Immunoglobulins and Intestinal Mucins.Biomolecules. 2021 Oct 31;11(11):1613. doi: 10.3390/biom11111613. Biomolecules. 2021. PMID: 34827611 Free PMC article.
-
α-Galactosidase and Sucrose-Kinase Relationships in a Bi-functional AgaSK Enzyme Produced by the Human Gut Symbiont Ruminococcus gnavus E1.Front Microbiol. 2020 Nov 12;11:579521. doi: 10.3389/fmicb.2020.579521. eCollection 2020. Front Microbiol. 2020. PMID: 33281771 Free PMC article.
-
Molecular advances in microbial α-galactosidases: challenges and prospects.World J Microbiol Biotechnol. 2022 Jul 1;38(9):148. doi: 10.1007/s11274-022-03340-2. World J Microbiol Biotechnol. 2022. PMID: 35773364 Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
