

The bone marrow-expressed antimicrobial cationic peptide LL-37 enhances the responsiveness of hematopoietic stem progenitor cells to an SDF-1 gradient and accelerates their engraftment after transplantation
- PMID: 21931324
- PMCID: PMC3244577
- DOI: 10.1038/leu.2011.252
The bone marrow-expressed antimicrobial cationic peptide LL-37 enhances the responsiveness of hematopoietic stem progenitor cells to an SDF-1 gradient and accelerates their engraftment after transplantation
Abstract
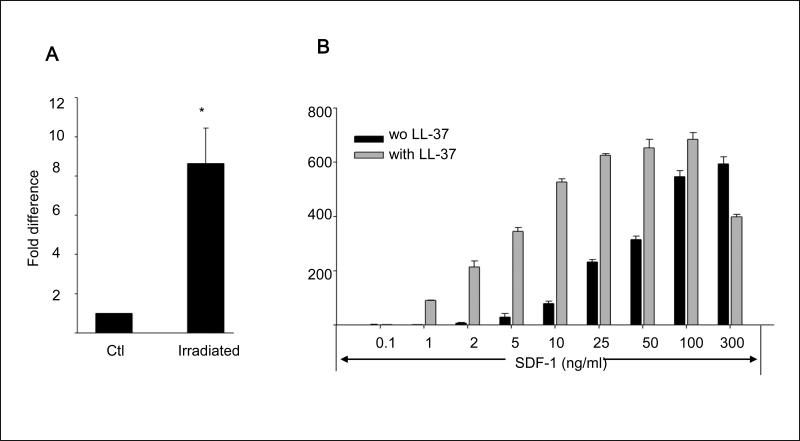
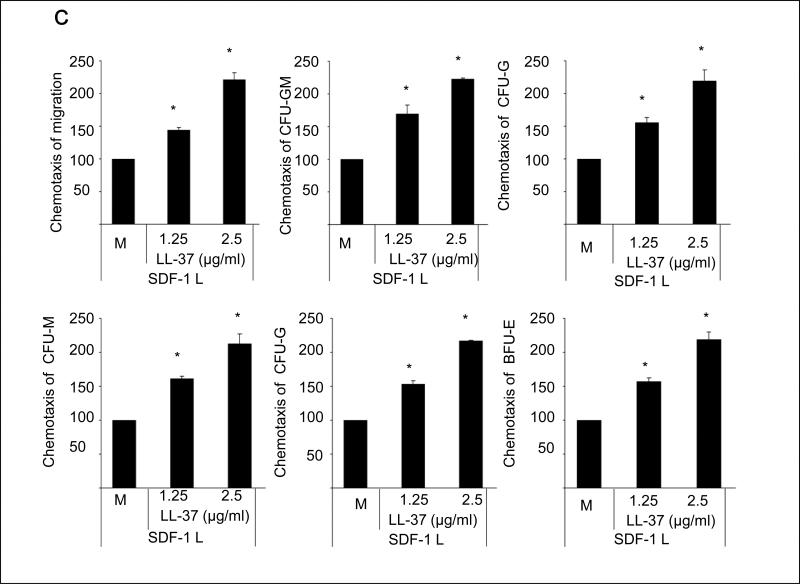
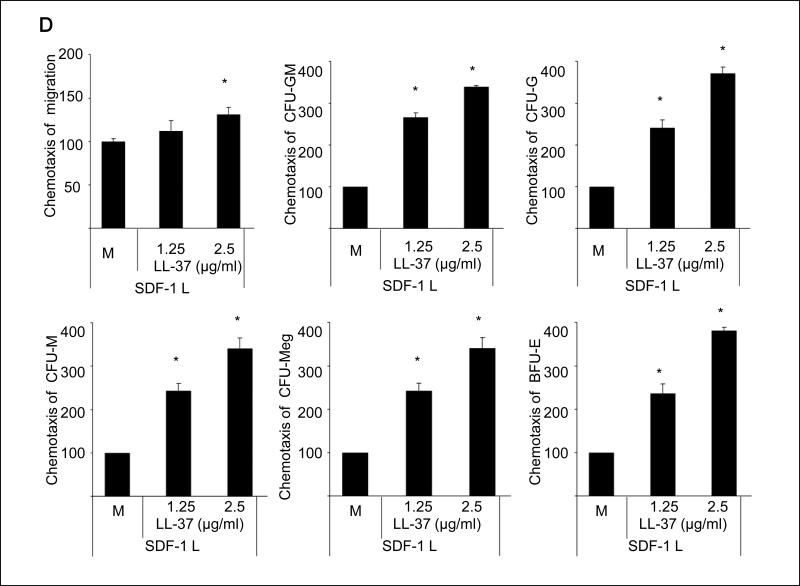
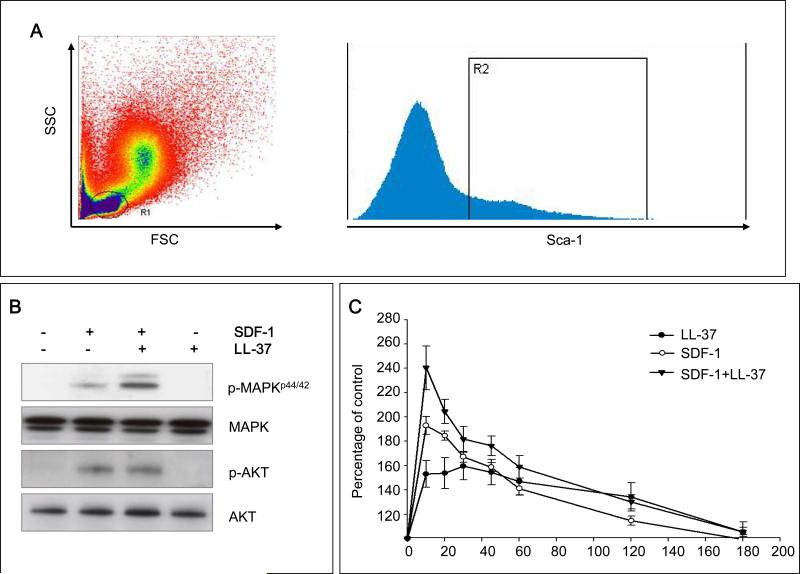
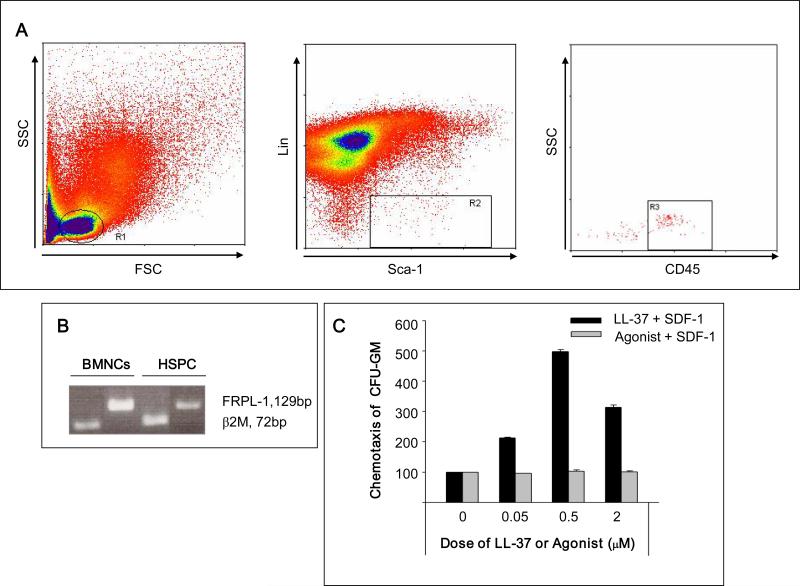
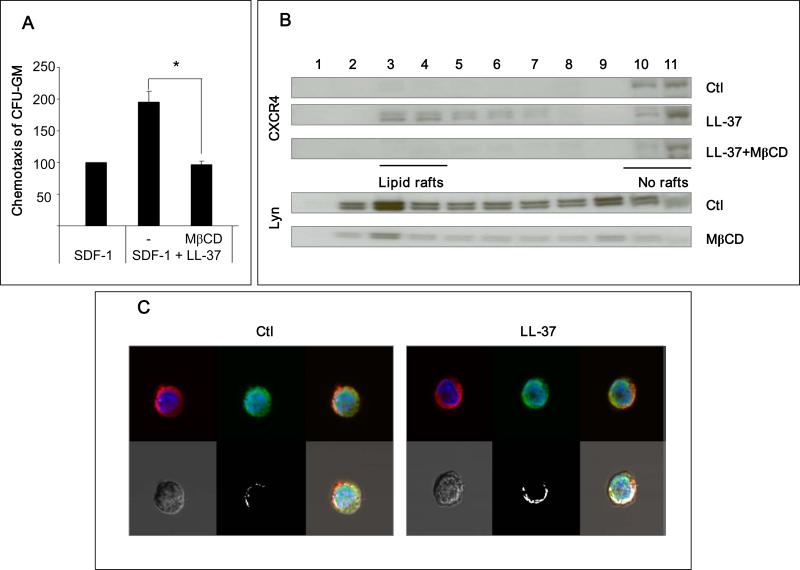
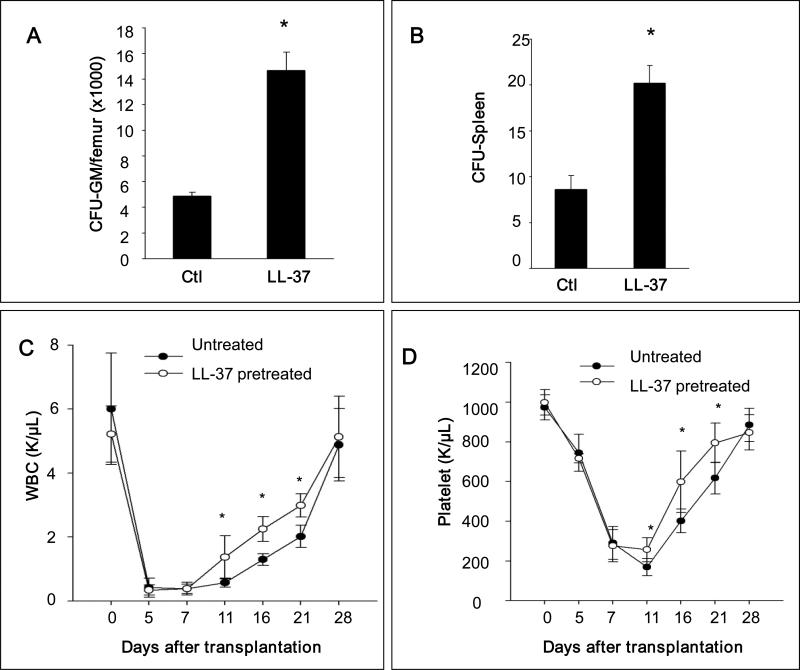
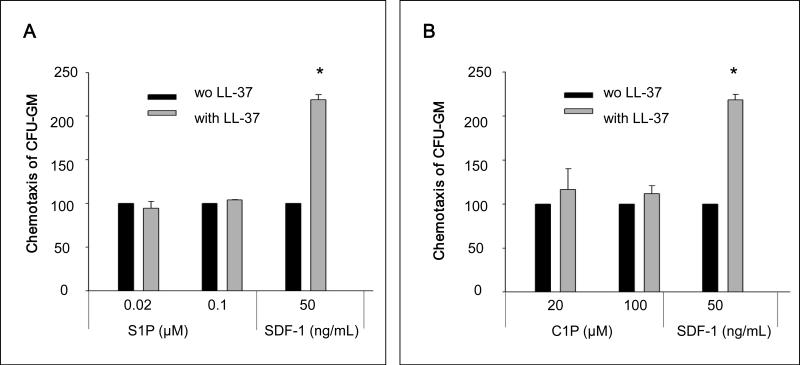
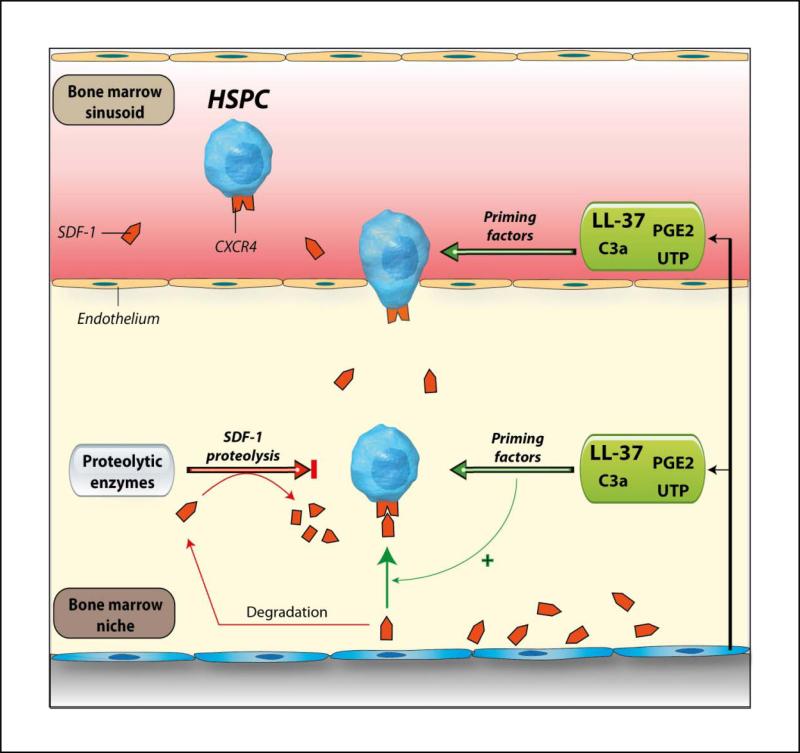
We report that the bone marrow (BM) stroma-released LL-37, a member of the cathelicidin family of antimicrobial peptides, primes/increases the responsiveness of murine and human hematopoietic stem/progenitor cells (HSPCs) to an α-chemokine stromal-derived factor-1 (SDF-1) gradient. Accordingly, LL-37 is upregulated in irradiated BM cells and enhances the chemotactic responsiveness of hematopoietic progenitors from all lineages to a low physiological SDF-1 gradient as well as increasing their (i) adhesiveness, (ii) SDF-1-mediated actin polymerization and (iii) MAPK(p42/44) phosphorylation. Mice transplanted with BM cells ex vivo primed by LL-37 showed accelerated recovery of platelet and neutrophil counts by ∼3-5 days compared with mice transplanted with unprimed control cells. These priming effects were not mediated by LL-37 binding to its receptor and depended instead on the incorporation of the CXCR4 receptor into membrane lipid rafts. We propose that LL-37, which has primarily antimicrobial functions and is harmless to mammalian cells, could be clinically applied to accelerate engraftment as an ex vivo priming agent for transplanted human HSPCs. This novel approach would be particularly important in cord blood transplantations, where the number of HSCs available is usually limited.
Figures
Similar articles
-
Incorporation of CXCR4 into membrane lipid rafts primes homing-related responses of hematopoietic stem/progenitor cells to an SDF-1 gradient.Blood. 2005 Jan 1;105(1):40-8. doi: 10.1182/blood-2004-04-1430. Epub 2004 Aug 24. Blood. 2005. PMID: 15328152
-
Innate immunity as orchestrator of bone marrow homing for hematopoietic stem/progenitor cells.Adv Exp Med Biol. 2013;735:219-32. doi: 10.1007/978-1-4614-4118-2_15. Adv Exp Med Biol. 2013. PMID: 23402030 Free PMC article. Review.
-
Dimethyl Sulfoxide (DMSO) Increases Percentage of CXCR4(+) Hematopoietic Stem/Progenitor Cells, Their Responsiveness to an SDF-1 Gradient, Homing Capacities, and Survival.Cell Transplant. 2016;25(7):1247-57. doi: 10.3727/096368915X689424. Epub 2015 Sep 4. Cell Transplant. 2016. PMID: 26345294
-
Nlrp3 Inflammasome Signaling Regulates the Homing and Engraftment of Hematopoietic Stem Cells (HSPCs) by Enhancing Incorporation of CXCR4 Receptor into Membrane Lipid Rafts.Stem Cell Rev Rep. 2020 Oct;16(5):954-967. doi: 10.1007/s12015-020-10005-w. Stem Cell Rev Rep. 2020. PMID: 32661868 Free PMC article.
-
Emerging Strategies to Enhance Homing and Engraftment of Hematopoietic Stem Cells.Stem Cell Rev Rep. 2016 Feb;12(1):121-8. doi: 10.1007/s12015-015-9625-5. Stem Cell Rev Rep. 2016. PMID: 26400757 Free PMC article. Review.
Cited by
-
A novel view of the adult bone marrow stem cell hierarchy and stem cell trafficking.Leukemia. 2015 Apr;29(4):776-82. doi: 10.1038/leu.2014.346. Epub 2014 Dec 9. Leukemia. 2015. PMID: 25486871 Free PMC article. Review.
-
Mouse Bone Marrow Sca-1+ CD44+ Mesenchymal Stem Cells Kill Avirulent Mycobacteria but Not Mycobacterium tuberculosis through Modulation of Cathelicidin Expression via the p38 Mitogen-Activated Protein Kinase-Dependent Pathway.Infect Immun. 2017 Sep 20;85(10):e00471-17. doi: 10.1128/IAI.00471-17. Print 2017 Oct. Infect Immun. 2017. PMID: 28739828 Free PMC article.
-
Further evidence that paroxysmal nocturnal haemoglobinuria is a disorder of defective cell membrane lipid rafts.J Cell Mol Med. 2015 Sep;19(9):2193-201. doi: 10.1111/jcmm.12605. Epub 2015 May 29. J Cell Mol Med. 2015. PMID: 26033571 Free PMC article.
-
Novel view on hematopoietic stem cell mobilization and homing.Leuk Suppl. 2014 Dec;3(Suppl 1):S19-20. doi: 10.1038/leusup.2014.11. Epub 2014 Dec 17. Leuk Suppl. 2014. PMID: 27175266 Free PMC article. No abstract available.
-
Downregulation of Heme Oxygenase 1 (HO-1) Activity in Hematopoietic Cells Enhances Their Engraftment After Transplantation.Cell Transplant. 2016;25(7):1265-76. doi: 10.3727/096368915X688957. Cell Transplant. 2016. PMID: 27412411 Free PMC article.
References
-
- Janowska-Wieczorek A, Majka M, Kijowski J, Baj-Krzyworzeka M, Reca R, Turner AR, et al. Platelet-derived microparticles bind to hematopoietic stem/progenitor cells and enhance their engraftment. Blood. 2001 Nov 15;98:3143–3149. 2001. - PubMed
-
- Reca R, Mastellos D, Majka M, Marquez L, Ratajczak J, Franchini S, et al. Functional receptor for C3a anaphylatoxin is expressed by normal hematopoietic stem/progenitor cells, and C3a enhances their homing-related responses to SDF-1. Blood. 2003 May 15;101:3784–3793. 2003. - PubMed
-
- Kimura T, Boehmler AM, Seitz G, Kuçi S, Wiesner T, Brinkmann V, et al. The sphingosine 1-phosphate receptor agonist FTY720 supports CXCR4-dependent migration and bone marrow homing of human CD34+ progenitor cells. Blood. 2004 Jun 15;103:4478–4486. 2004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
