

The hedgehog receptor patched is involved in cholesterol transport
- PMID: 21931618
- PMCID: PMC3169562
- DOI: 10.1371/journal.pone.0023834
The hedgehog receptor patched is involved in cholesterol transport
Abstract
Background: Sonic hedgehog (Shh) signaling plays a crucial role in growth and patterning during embryonic development, and also in stem cell maintenance and tissue regeneration in adults. Aberrant Shh pathway activation is involved in the development of many tumors, and one of the most affected Shh signaling steps found in these tumors is the regulation of the signaling receptor Smoothened by the Shh receptor Patched. In the present work, we investigated Patched activity and the mechanism by which Patched inhibits Smoothened.
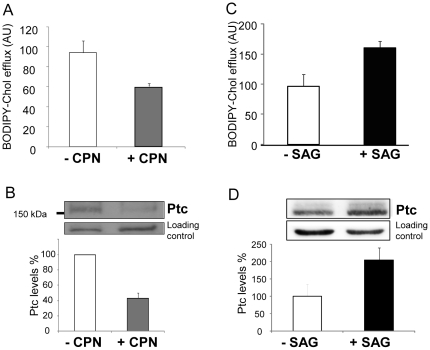
Methodology/principal findings: Using the well-known Shh-responding cell line of mouse fibroblasts NIH 3T3, we first observed that enhancement of the intracellular cholesterol concentration induces Smoothened enrichment in the plasma membrane, which is a crucial step for the signaling activation. We found that binding of Shh protein to its receptor Patched, which involves Patched internalization, increases the intracellular concentration of cholesterol and decreases the efflux of a fluorescent cholesterol derivative (BODIPY-cholesterol) from these cells. Treatment of fibroblasts with cyclopamine, an antagonist of Shh signaling, inhibits Patched expression and reduces BODIPY-cholesterol efflux, while treatment with the Shh pathway agonist SAG enhances Patched protein expression and BODIPY-cholesterol efflux. We also show that over-expression of human Patched in the yeast S. cerevisiae results in a significant boost of BODIPY-cholesterol efflux. Furthermore, we demonstrate that purified Patched binds to cholesterol, and that the interaction of Shh with Patched inhibits the binding of Patched to cholesterol.
Conclusion/significance: Our results suggest that Patched may contribute to cholesterol efflux from cells, and to modulation of the intracellular cholesterol concentration. This activity is likely responsible for the inhibition of the enrichment of Smoothened in the plasma membrane, which is an important step in Shh pathway activation.
Conflict of interest statement
Figures





References
-
- Varjosalo M, Taipale J. Hedgehog: functions and mechanisms. Genes Dev. 2008;22:2454–2472. - PubMed
-
- Bale A. Hedgehog signaling and human disease. Annu Rev Genomics Hum Genet. 2002;3:47–65. - PubMed
-
- Nieuwenhuis E, Hui C. Hedgehog signaling and congenital malformations. Clin Genet. 2005;67:193–208. - PubMed
-
- Nehmé R, Mus-Veteau I. Proteins of the Hedgehog signaling pathway as therapeutic targets against cancer. Expert Rev Proteomics. 2010;7:601–612. - PubMed
-
- Beachy P, Karhadkar S, Berman D. Tissue repair and stem cell renewal in carcinogenesis. Nature. 2004;432:324–331. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
