

Human embryonic and fetal mesenchymal stem cells differentiate toward three different cardiac lineages in contrast to their adult counterparts
- PMID: 21931658
- PMCID: PMC3170333
- DOI: 10.1371/journal.pone.0024164
Human embryonic and fetal mesenchymal stem cells differentiate toward three different cardiac lineages in contrast to their adult counterparts
Abstract
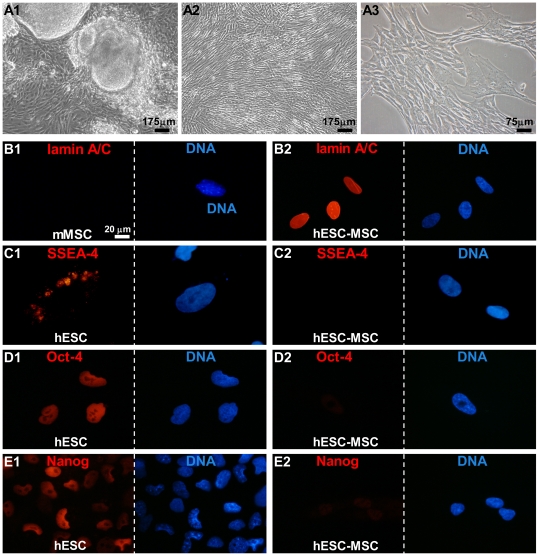
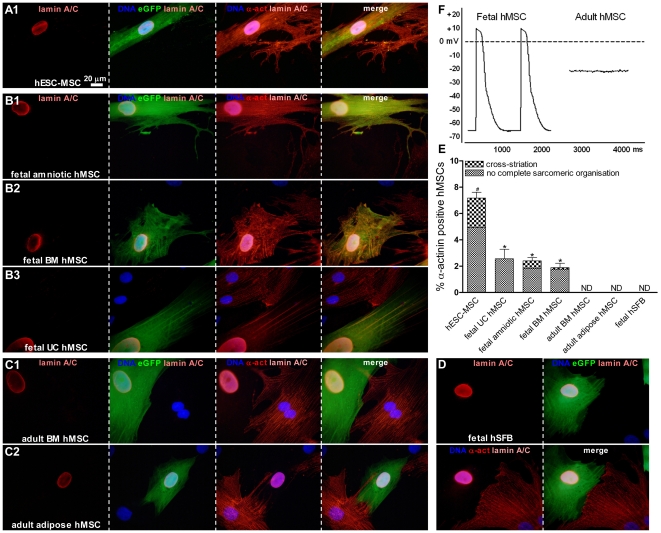
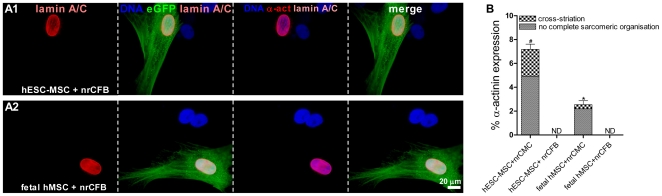
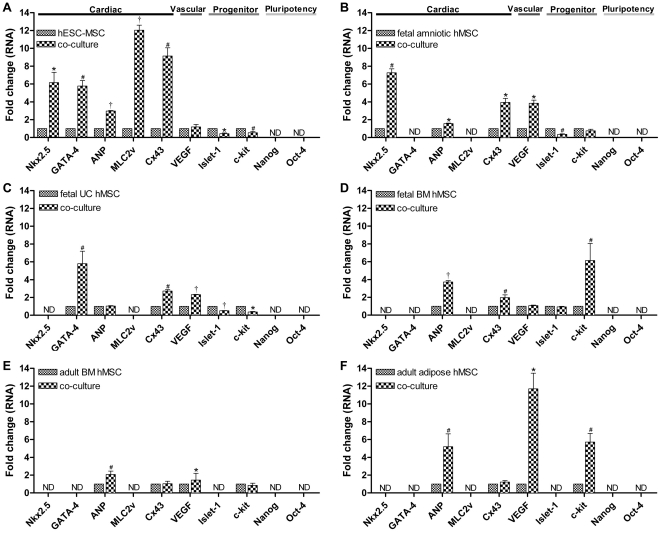
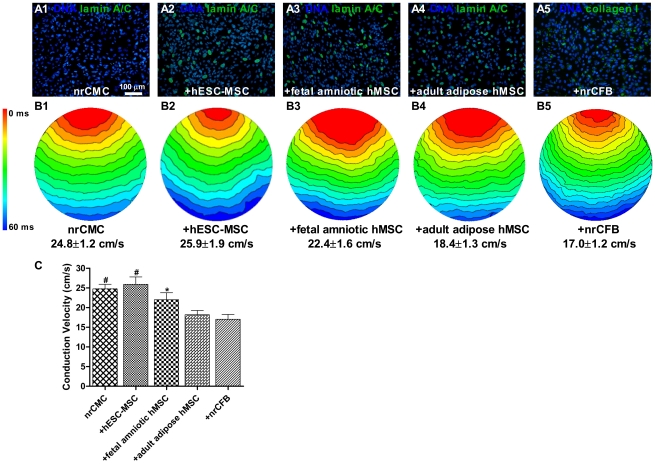
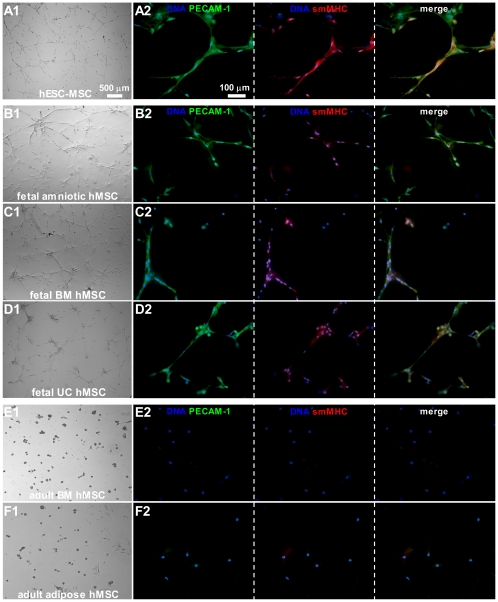
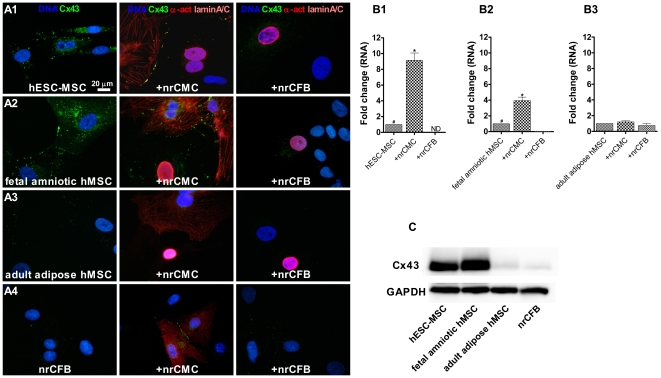
Mesenchymal stem cells (MSCs) show unexplained differences in differentiation potential. In this study, differentiation of human (h) MSCs derived from embryonic, fetal and adult sources toward cardiomyocytes, endothelial and smooth muscle cells was investigated. Labeled hMSCs derived from embryonic stem cells (hESC-MSCs), fetal umbilical cord, bone marrow, amniotic membrane and adult bone marrow and adipose tissue were co-cultured with neonatal rat cardiomyocytes (nrCMCs) or cardiac fibroblasts (nrCFBs) for 10 days, and also cultured under angiogenic conditions. Cardiomyogenesis was assessed by human-specific immunocytological analysis, whole-cell current-clamp recordings, human-specific qRT-PCR and optical mapping. After co-culture with nrCMCs, significantly more hESC-MSCs than fetal hMSCs stained positive for α-actinin, whereas adult hMSCs stained negative. Furthermore, functional cardiomyogenic differentiation, based on action potential recordings, was shown to occur, but not in adult hMSCs. Of all sources, hESC-MSCs expressed most cardiac-specific genes. hESC-MSCs and fetal hMSCs contained significantly higher basal levels of connexin43 than adult hMSCs and co-culture with nrCMCs increased expression. After co-culture with nrCFBs, hESC-MSCs and fetal hMSCs did not express α-actinin and connexin43 expression was decreased. Conduction velocity (CV) in co-cultures of nrCMCs and hESC-MSCs was significantly higher than in co-cultures with fetal or adult hMSCs. In angiogenesis bioassays, only hESC-MSCs and fetal hMSCs were able to form capillary-like structures, which stained for smooth muscle and endothelial cell markers.Human embryonic and fetal MSCs differentiate toward three different cardiac lineages, in contrast to adult MSCs. Cardiomyogenesis is determined by stimuli from the cellular microenvironment, where connexin43 may play an important role.
Conflict of interest statement
Figures
Similar articles
-
Gap junctional coupling with cardiomyocytes is necessary but not sufficient for cardiomyogenic differentiation of cocultured human mesenchymal stem cells.Stem Cells. 2012 Jun;30(6):1236-45. doi: 10.1002/stem.1086. Stem Cells. 2012. PMID: 22438316
-
Bone marrow-derived human mesenchymal stem cells express cardiomyogenic proteins but do not exhibit functional cardiomyogenic differentiation potential.Stem Cells Dev. 2012 Sep 1;21(13):2457-70. doi: 10.1089/scd.2011.0626. Epub 2012 Mar 13. Stem Cells Dev. 2012. PMID: 22309203 Free PMC article.
-
Different cardiovascular potential of adult- and fetal-type mesenchymal stem cells in a rat model of heart cryoinjury.Cell Transplant. 2008;17(6):679-94. doi: 10.3727/096368908786092739. Cell Transplant. 2008. PMID: 18819256
-
Human embryonic stem cell-derived mesenchymal progenitors: an overview.Methods Mol Biol. 2011;690:163-74. doi: 10.1007/978-1-60761-962-8_11. Methods Mol Biol. 2011. PMID: 21042992 Review.
-
Human placental stem cells: biomedical potential and clinical relevance.J Stem Cells. 2011;6(2):75-92. J Stem Cells. 2011. PMID: 22997848 Review.
Cited by
-
Human embryonic stem cell derived mesenchymal progenitors express cardiac markers but do not form contractile cardiomyocytes.PLoS One. 2013;8(1):e54524. doi: 10.1371/journal.pone.0054524. Epub 2013 Jan 16. PLoS One. 2013. PMID: 23342164 Free PMC article.
-
Donor mesenchymal stromal cells (MSCs) undergo variable cardiac reprogramming in vivo and predominantly co-express cardiac and stromal determinants after experimental acute myocardial infarction.Stem Cell Rev Rep. 2014 Apr;10(2):304-15. doi: 10.1007/s12015-013-9483-y. Stem Cell Rev Rep. 2014. PMID: 24287730
-
High incidence of contaminating maternal cell overgrowth in human placental mesenchymal stem/stromal cell cultures: a systematic review.Stem Cells Transl Med. 2014 Nov;3(11):1305-11. doi: 10.5966/sctm.2014-0051. Epub 2014 Aug 25. Stem Cells Transl Med. 2014. PMID: 25154781 Free PMC article.
-
Sublethal heat shock induces premature senescence rather than apoptosis in human mesenchymal stem cells.Cell Stress Chaperones. 2014 May;19(3):355-66. doi: 10.1007/s12192-013-0463-6. Cell Stress Chaperones. 2014. PMID: 24078383 Free PMC article.
-
Cancer-associated fibroblast-derived Gremlin 1 promotes breast cancer progression.Breast Cancer Res. 2019 Sep 18;21(1):109. doi: 10.1186/s13058-019-1194-0. Breast Cancer Res. 2019. PMID: 31533776 Free PMC article.
References
-
- Lloyd-Jones D, Adams R, Carnethon M, De SG, Ferguson TB, et al. Heart disease and stroke statistics–2009 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation. 2009;119:480–486. - PubMed
-
- Ballard VL, Edelberg JM. Stem cells and the regeneration of the aging cardiovascular system. Circ Res. 2007;100:1116–1127. - PubMed
-
- Passier R, van Laake LW, Mummery CL. Stem-cell-based therapy and lessons from the heart. Nature. 2008;453:322–329. - PubMed
-
- O'Donoghue K, Fisk NM. Fetal stem cells. Best Pract Res Clin Obstet Gynaecol. 2004;18:853–875. - PubMed
-
- Roobrouck VD, Ulloa-Montoya F, Verfaillie CM. Self-renewal and differentiation capacity of young and aged stem cells. Exp Cell Res. 2008;314:1937–1944. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
