

Morphological changes in diabetic kidney are associated with increased O-GlcNAcylation of cytoskeletal proteins including α-actinin 4
- PMID: 21933451
- PMCID: PMC3224550
- DOI: 10.1186/1559-0275-8-15
Morphological changes in diabetic kidney are associated with increased O-GlcNAcylation of cytoskeletal proteins including α-actinin 4
Abstract
Purpose: The objective of the present study is to identify proteins that change in the extent of the modification with O-linked N-acetylglucosamine (O-GlcNAcylation) in the kidney from diabetic model Goto-Kakizaki (GK) rats, and to discuss the relation between O-GlcNAcylation and the pathological condition in diabetes.
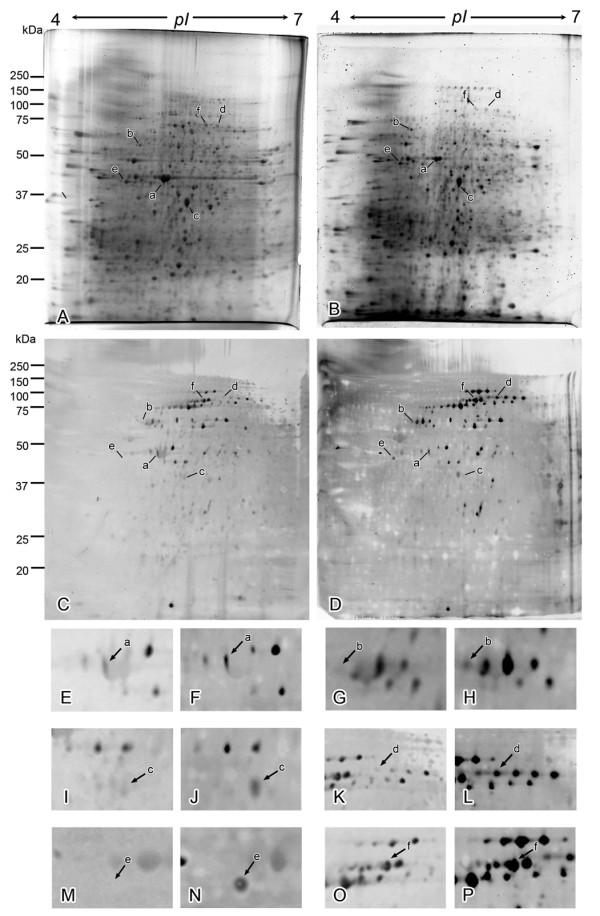
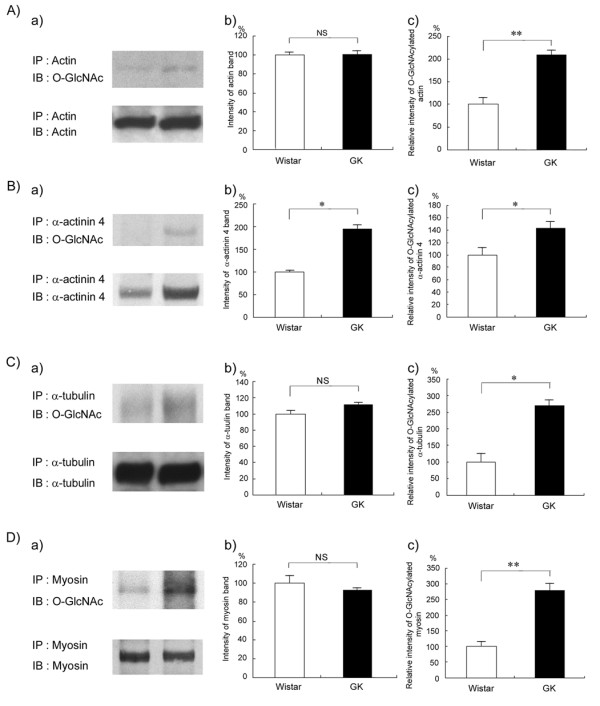
Methods: O-GlcNAcylated proteins were identified by two-dimensional gel electrophoresis, immunoblotting and peptide mass fingerprinting. The level of O-GlcNAcylation of these proteins was examined by immunoprecipitation, immunoblotting and in situ Proximity Ligation Assay (PLA).
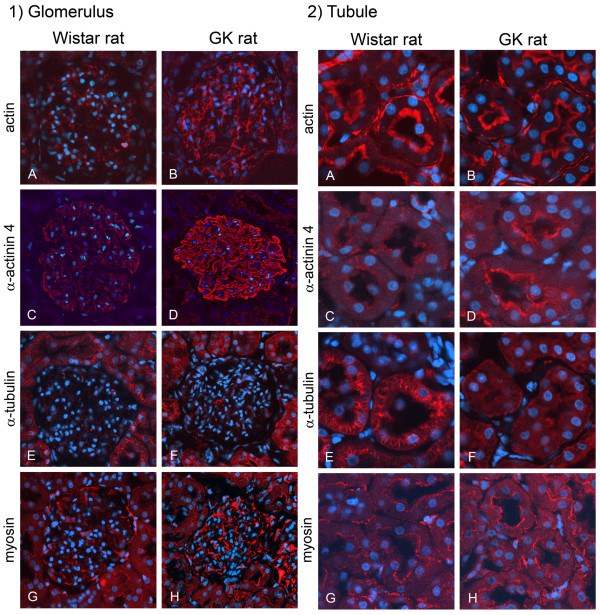
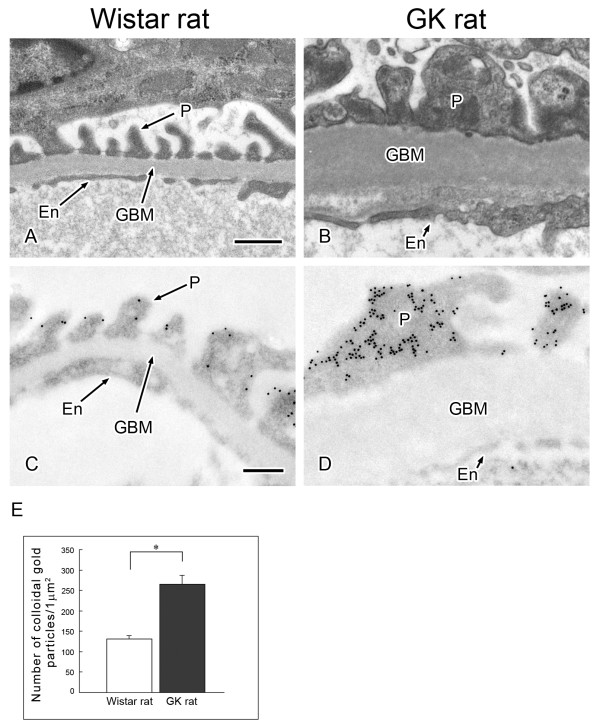
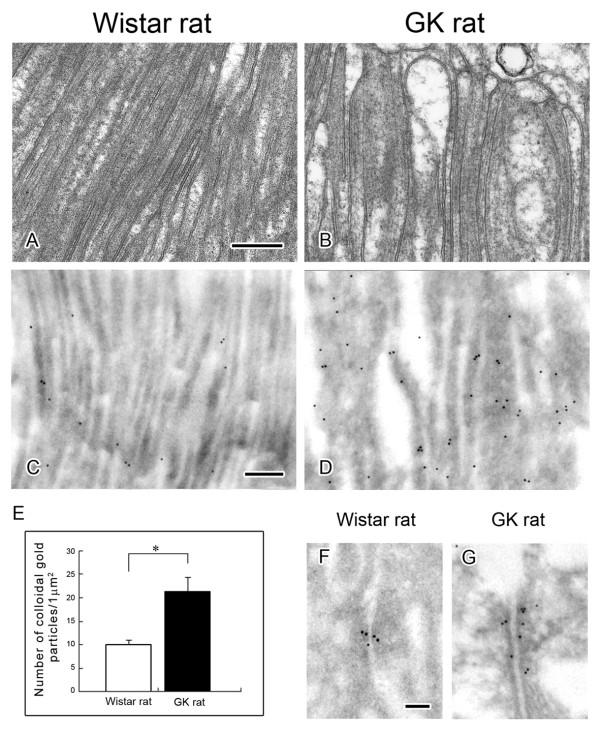
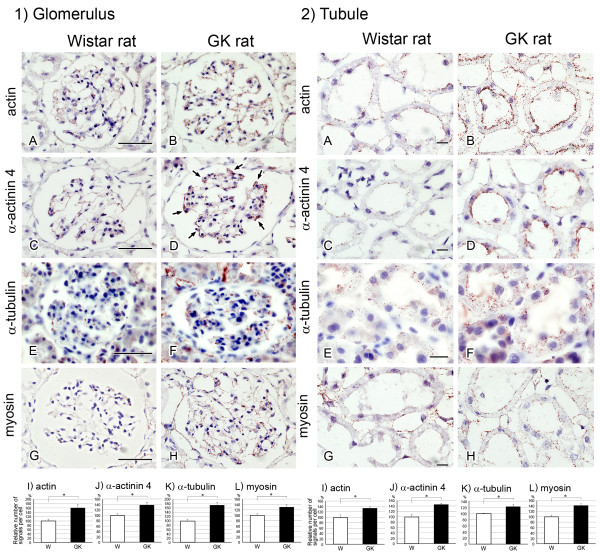
Results: O-GlcNAcylated proteins that changed significantly in the degree of O-GlcNAcylation were identified as cytoskeletal proteins (α-actin, α-tubulin, α-actinin 4, myosin) and mitochondrial proteins (ATP synthase β, pyruvate carboxylase). The extent of O-GlcNAcylation of the above proteins increased in the diabetic kidney. Immunofluorescence and in situ PLA studies revealed that the levels of O-GlcNAcylation of actin, α-actinin 4 and myosin were significantly increased in the glomerulus and the proximal tubule of the diabetic kidney. Immunoelectron microscopy revealed that immunolabeling of α-actinin 4 is disturbed and increased in the foot process of podocytes of glomerulus and in the microvilli of proximal tubules.
Conclusion: These results suggest that changes in the O-GlcNAcylation of cytoskeletal proteins are closely associated with the morphological changes in the podocyte foot processes in the glomerulus and in microvilli of proximal tubules in the diabetic kidney. This is the first report to show that α-actinin 4 is O-GlcNAcylated. α-Actinin 4 will be a good marker protein to examine the relation between O-GlcNAcylation and diabetic nephropathy.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases