

Nonessential plastid-encoded ribosomal proteins in tobacco: a developmental role for plastid translation and implications for reductive genome evolution
- PMID: 21934145
- PMCID: PMC3203423
- DOI: 10.1105/tpc.111.088906
Nonessential plastid-encoded ribosomal proteins in tobacco: a developmental role for plastid translation and implications for reductive genome evolution
Abstract
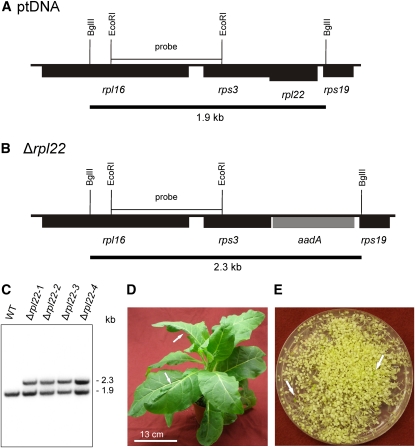
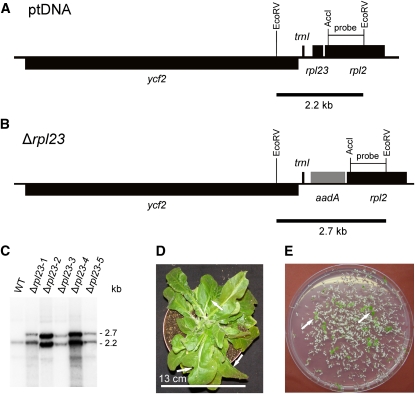
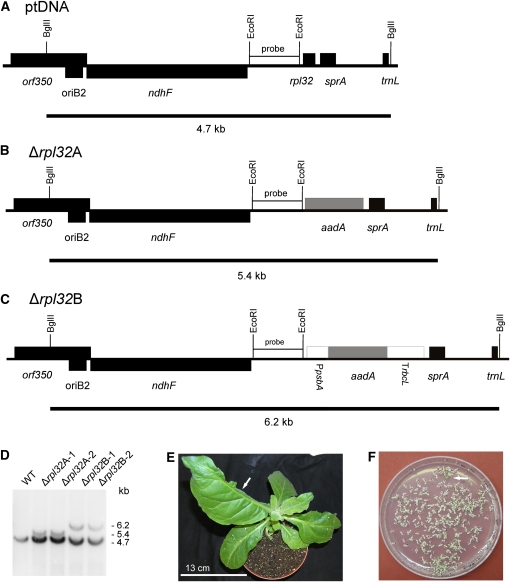
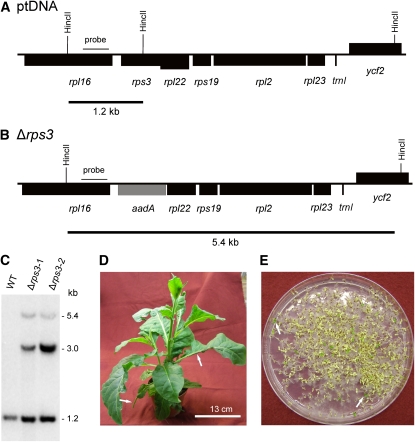
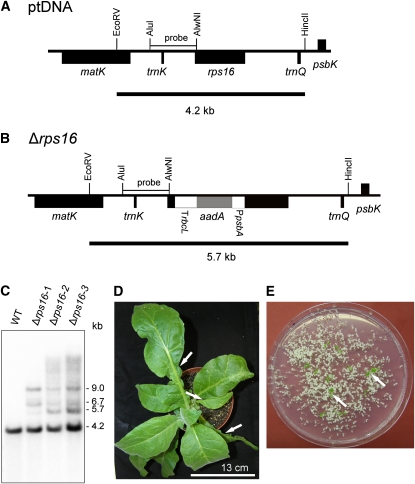
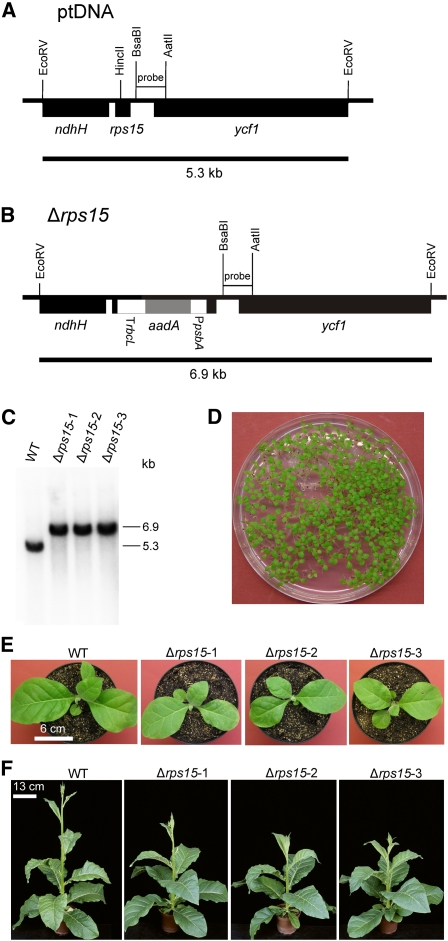
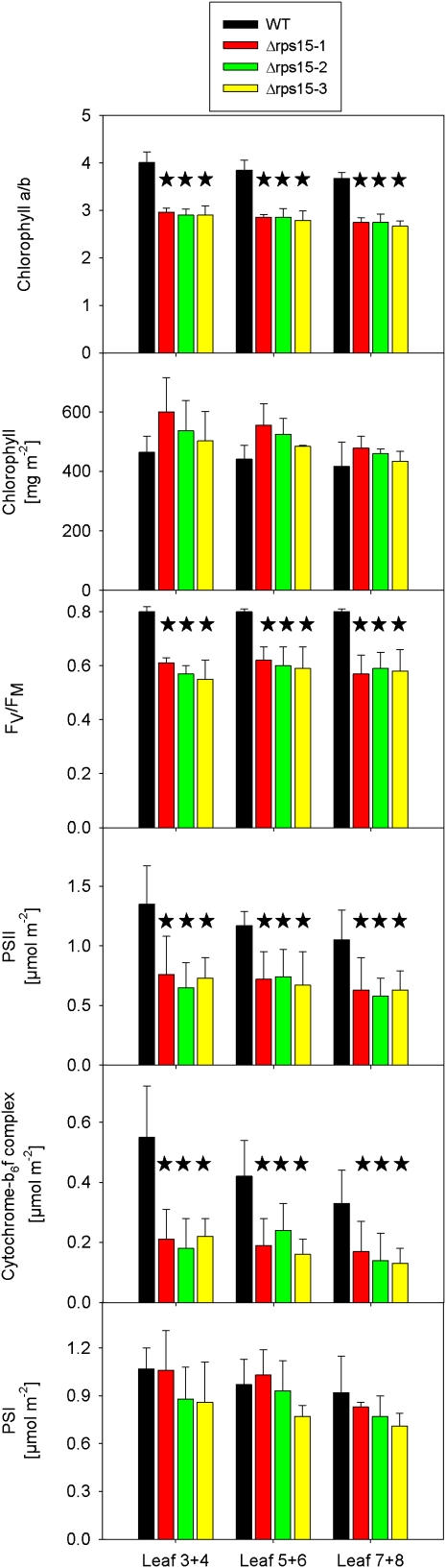
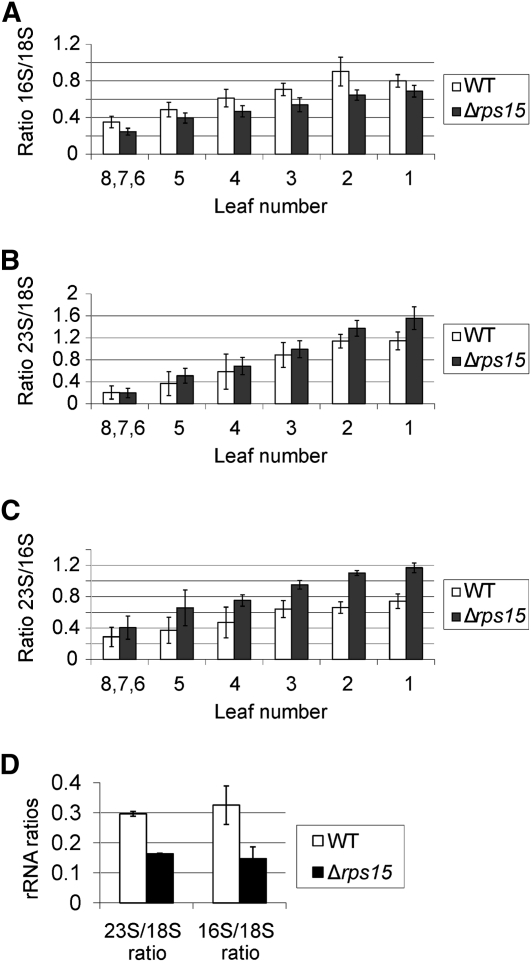
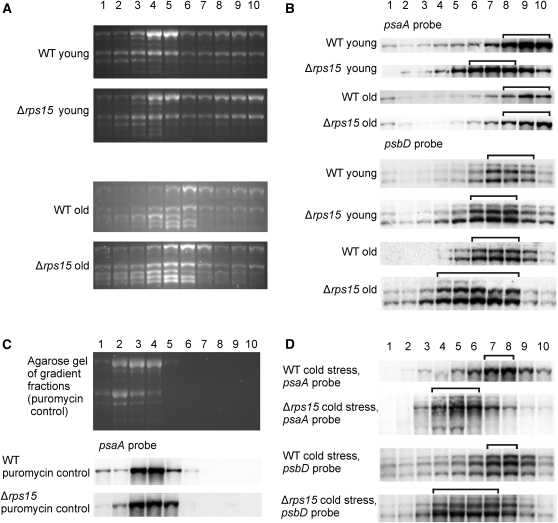
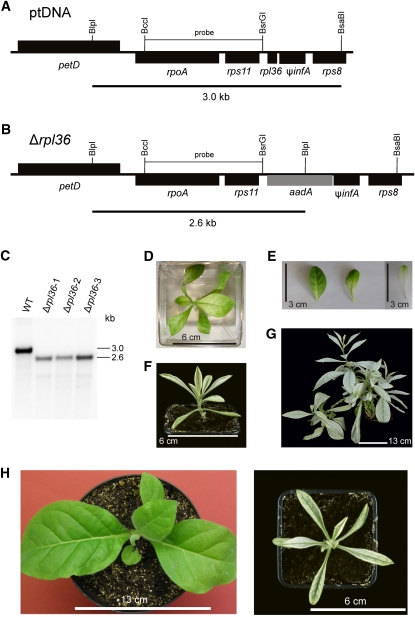
Plastid genomes of higher plants contain a conserved set of ribosomal protein genes. Although plastid translational activity is essential for cell survival in tobacco (Nicotiana tabacum), individual plastid ribosomal proteins can be nonessential. Candidates for nonessential plastid ribosomal proteins are ribosomal proteins identified as nonessential in bacteria and those whose genes were lost from the highly reduced plastid genomes of nonphotosynthetic plastid-bearing lineages (parasitic plants, apicomplexan protozoa). Here we report the reverse genetic analysis of seven plastid-encoded ribosomal proteins that meet these criteria. We have introduced knockout alleles for the corresponding genes into the tobacco plastid genome. Five of the targeted genes (ribosomal protein of the large subunit22 [rpl22], rpl23, rpl32, ribosomal protein of the small subunit3 [rps3], and rps16) were shown to be essential even under heterotrophic conditions, despite their loss in at least some parasitic plastid-bearing lineages. This suggests that nonphotosynthetic plastids show elevated rates of gene transfer to the nuclear genome. Knockout of two ribosomal protein genes, rps15 and rpl36, yielded homoplasmic transplastomic mutants, thus indicating nonessentiality. Whereas Δrps15 plants showed only a mild phenotype, Δrpl36 plants were severely impaired in photosynthesis and growth and, moreover, displayed greatly altered leaf morphology. This finding provides strong genetic evidence that chloroplast translational activity influences leaf development, presumably via a retrograde signaling pathway.
Figures

Similar articles
-
Rpl33, a nonessential plastid-encoded ribosomal protein in tobacco, is required under cold stress conditions.Plant Cell. 2008 Aug;20(8):2221-37. doi: 10.1105/tpc.108.060392. Epub 2008 Aug 29. Plant Cell. 2008. PMID: 18757552 Free PMC article.
-
Synthetic lethality in the tobacco plastid ribosome and its rescue at elevated growth temperatures.Plant Cell. 2014 Feb;26(2):765-76. doi: 10.1105/tpc.114.123240. Epub 2014 Feb 21. Plant Cell. 2014. PMID: 24563204 Free PMC article.
-
The plastid-specific ribosomal proteins of Arabidopsis thaliana can be divided into non-essential proteins and genuine ribosomal proteins.Plant J. 2012 Jan;69(2):302-16. doi: 10.1111/j.1365-313X.2011.04791.x. Epub 2011 Oct 21. Plant J. 2012. PMID: 21923745
-
The translational apparatus of plastids and its role in plant development.Mol Plant. 2014 Jul;7(7):1105-20. doi: 10.1093/mp/ssu022. Epub 2014 Mar 3. Mol Plant. 2014. PMID: 24589494 Free PMC article. Review.
-
Unveiling the functions of plastid ribosomal proteins in plant development and abiotic stress tolerance.Plant Physiol Biochem. 2022 Oct 15;189:35-45. doi: 10.1016/j.plaphy.2022.07.029. Epub 2022 Aug 2. Plant Physiol Biochem. 2022. PMID: 36041366 Review.
Cited by
-
The Arabidopsis organelle-localized glycyl-tRNA synthetase encoded by EMBRYO DEFECTIVE DEVELOPMENT1 is required for organ patterning.J Exp Bot. 2012 Sep;63(14):5233-43. doi: 10.1093/jxb/ers184. Epub 2012 Jul 12. J Exp Bot. 2012. PMID: 22791832 Free PMC article.
-
Comparative Plastome Analysis of Root- and Stem-Feeding Parasites of Santalales Untangle the Footprints of Feeding Mode and Lifestyle Transitions.Genome Biol Evol. 2020 Jan 1;12(1):3663-3676. doi: 10.1093/gbe/evz271. Genome Biol Evol. 2020. PMID: 31845987 Free PMC article.
-
Variations and reduction of plastome are associated with the evolution of parasitism in Convolvulaceae.Plant Mol Biol. 2024 Apr 15;114(3):40. doi: 10.1007/s11103-024-01440-1. Plant Mol Biol. 2024. PMID: 38622367
-
Identification of enzymes for adenosine-to-inosine editing and discovery of cytidine-to-uridine editing in nucleus-encoded transfer RNAs of Arabidopsis.Plant Physiol. 2014 Dec;166(4):1985-97. doi: 10.1104/pp.114.250498. Epub 2014 Oct 14. Plant Physiol. 2014. PMID: 25315605 Free PMC article.
-
Comparative chloroplast genome analyses of potentilleae: insights into genome characteristics, mutational hotspots, and adaptive evolution.Genetica. 2025 Jun 10;153(1):22. doi: 10.1007/s10709-025-00236-5. Genetica. 2025. PMID: 40493127
References
-
- Albrecht V., Ingenfeld A., Apel K. (2006). Characterization of the snowy cotyledon 1 mutant of Arabidopsis thaliana: The impact of chloroplast elongation factor G on chloroplast development and plant vitality. Plant Mol. Biol. 60: 507–518 - PubMed
-
- Barbrook A.C., Howe C.J., Purton S. (2006). Why are plastid genomes retained in nonphotosynthetic organisms? Trends Plant Sci. 11: 101–108 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
