

Hippocampal neurogenesis and dendritic plasticity support running-improved spatial learning and depression-like behaviour in stressed rats
- PMID: 21935393
- PMCID: PMC3174166
- DOI: 10.1371/journal.pone.0024263
Hippocampal neurogenesis and dendritic plasticity support running-improved spatial learning and depression-like behaviour in stressed rats
Abstract
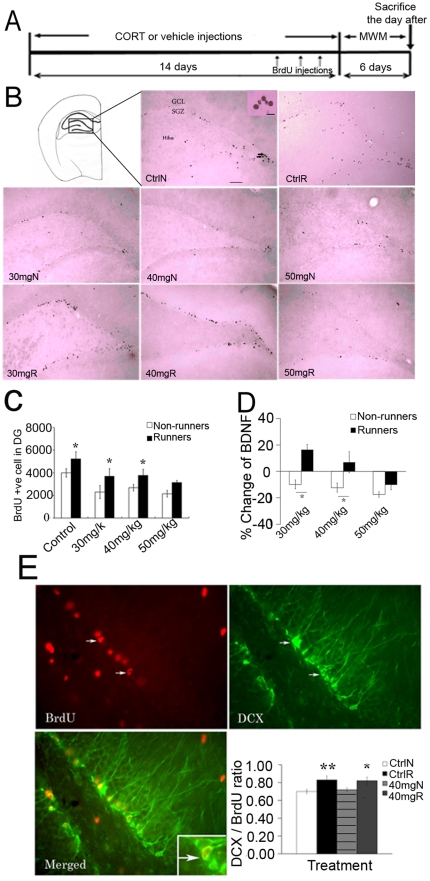
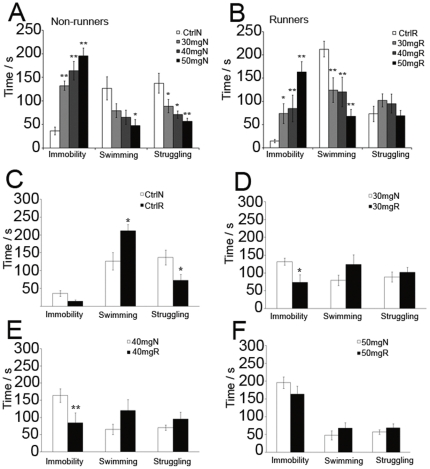
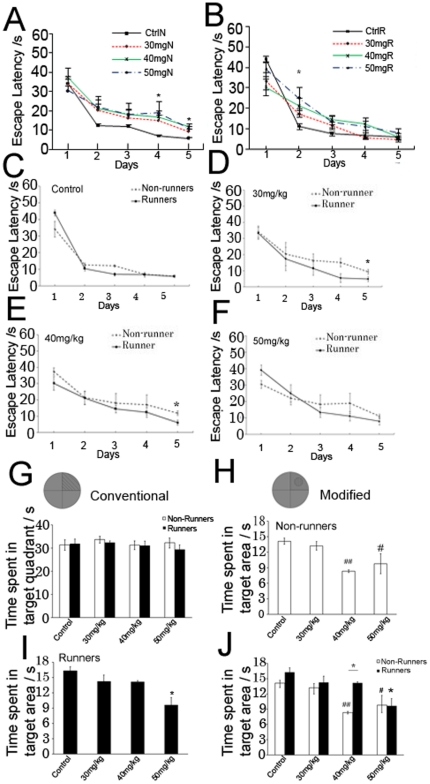
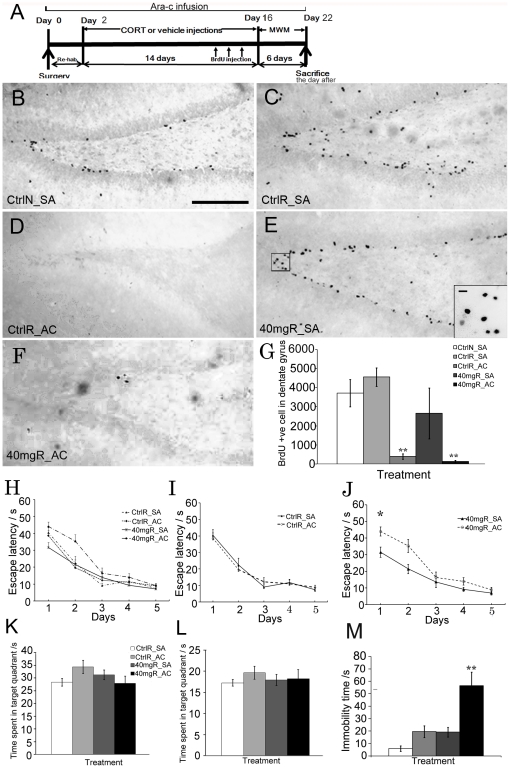
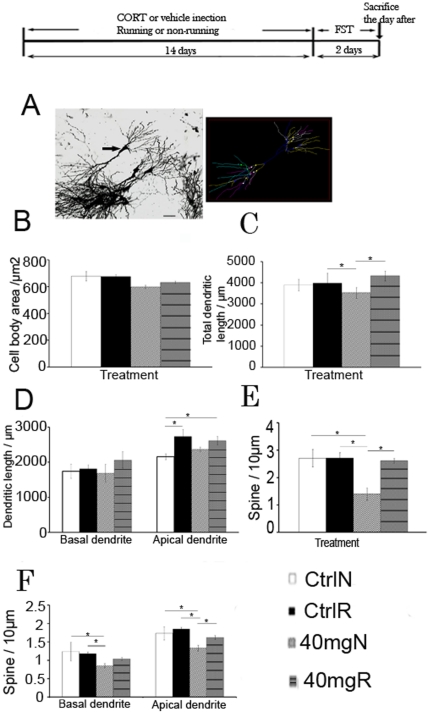
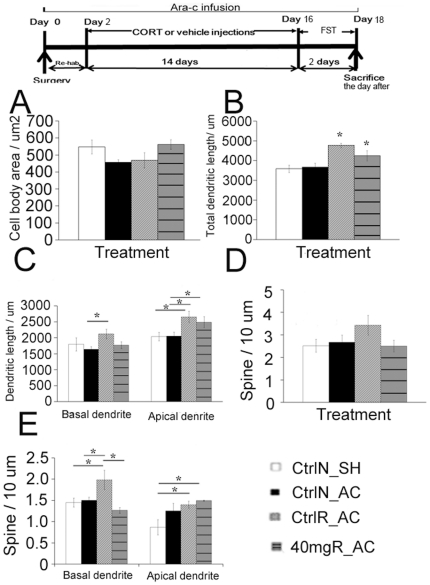
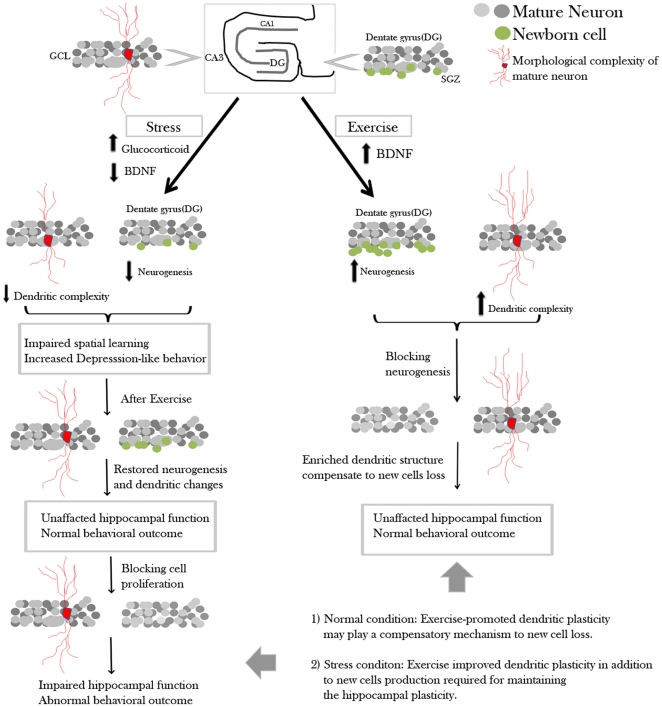
Exercise promotes hippocampal neurogenesis and dendritic plasticity while stress shows the opposite effects, suggesting a possible mechanism for exercise to counteract stress. Changes in hippocampal neurogenesis and dendritic modification occur simultaneously in rats with stress or exercise; however, it is unclear whether neurogenesis or dendritic remodeling has a greater impact on mediating the effect of exercise on stress since they have been separately examined. Here we examined hippocampal cell proliferation in runners treated with different doses (low: 30 mg/kg; moderate: 40 mg/kg; high: 50 mg/kg) of corticosterone (CORT) for 14 days. Water maze task and forced swim tests were applied to assess hippocampal-dependent learning and depression-like behaviour respectively the day after the treatment. Repeated CORT treatment resulted in a graded increase in depression-like behaviour and impaired spatial learning that is associated with decreased hippocampal cell proliferation and BDNF levels. Running reversed these effects in rats treated with low or moderate, but not high doses of CORT. Using 40 mg/kg CORT-treated rats, we further studied the role of neurogenesis and dendritic remodeling in mediating the effects of exercise on stress. Co-labelling with BrdU (thymidine analog) /doublecortin (immature neuronal marker) showed that running increased neuronal differentiation in vehicle- and CORT-treated rats. Running also increased dendritic length and spine density in CA3 pyramidal neurons in 40 mg/kg CORT-treated rats. Ablation of neurogenesis with Ara-c infusion diminished the effect of running on restoring spatial learning and decreasing depression-like behaviour in 40 mg/kg CORT-treated animals in spite of dendritic and spine enhancement. but not normal runners with enhanced dendritic length. The results indicate that both restored hippocampal neurogenesis and dendritic remodelling within the hippocampus are essential for running to counteract stress.
Conflict of interest statement
Figures
References
-
- Redila VA, Christie BR. Exercise-induced changes in dendritic structure and complexity in the adult hippocampal dentate gyrus. Neuroscience. 2006;137:1299–1307. - PubMed
-
- Neeper SA, Gomez-Pinilla F, Choi J, Cotman CW. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res. 1996;726:49–56. - PubMed
-
- Watanabe Y, Gould E, McEwen BS. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res. 1992;588:341–345. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
