

A rooted net of life
- PMID: 21936906
- PMCID: PMC3189188
- DOI: 10.1186/1745-6150-6-45
A rooted net of life
Abstract
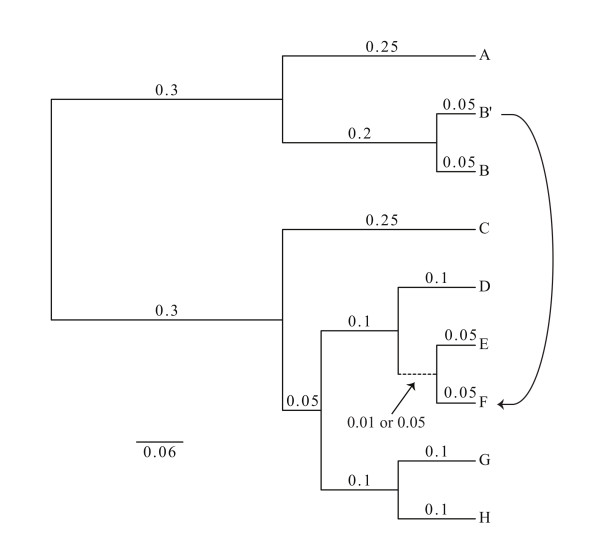
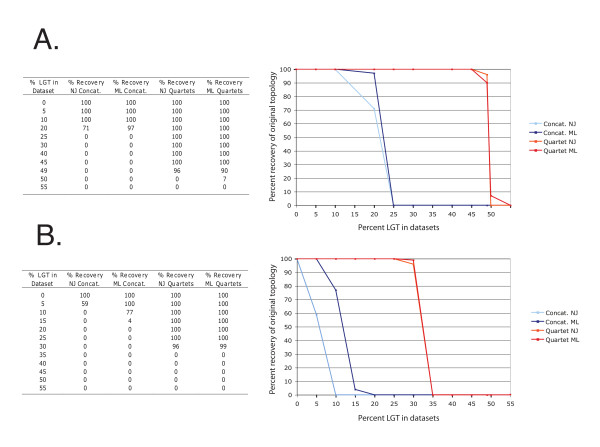
Phylogenetic reconstruction using DNA and protein sequences has allowed the reconstruction of evolutionary histories encompassing all life. We present and discuss a means to incorporate much of this rich narrative into a single model that acknowledges the discrete evolutionary units that constitute the organism. Briefly, this Rooted Net of Life genome phylogeny is constructed around an initial, well resolved and rooted tree scaffold inferred from a supermatrix of combined ribosomal genes. Extant sampled ribosomes form the leaves of the tree scaffold. These leaves, but not necessarily the deeper parts of the scaffold, can be considered to represent a genome or pan-genome, and to be associated with members of other gene families within that sequenced (pan)genome. Unrooted phylogenies of gene families containing four or more members are reconstructed and superimposed over the scaffold. Initially, reticulations are formed where incongruities between topologies exist. Given sufficient evidence, edges may then be differentiated as those representing vertical lines of inheritance within lineages and those representing horizontal genetic transfers or endosymbioses between lineages.
Figures
References
-
- Wu DY, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Ivanova NN, Kunin V, Goodwin L, Wu M, Tindall BJ, Hooper SD, Pati A, Lykidis A, Spring S, Anderson IJ, D'haeseleer P, Zemla A, Singer M, Lapidus A, Nolan M, Copeland A, Han C, Chen F, Cheng JF, Lucas S, Kerfeld C, Lang E, Gronow S, Chain P, Bruce D, Rubin EM, Kyrpides NC, Klenk HP, Eisen JA. A phylogeny-driven genomic encyclopaedia of Bacteria and Archaea. Nature. 2009;462(7276):1056–1060. doi: 10.1038/nature08656. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
