

Human telomerase acts as a hTR-independent reverse transcriptase in mitochondria
- PMID: 21937513
- PMCID: PMC3258147
- DOI: 10.1093/nar/gkr758
Human telomerase acts as a hTR-independent reverse transcriptase in mitochondria
Abstract
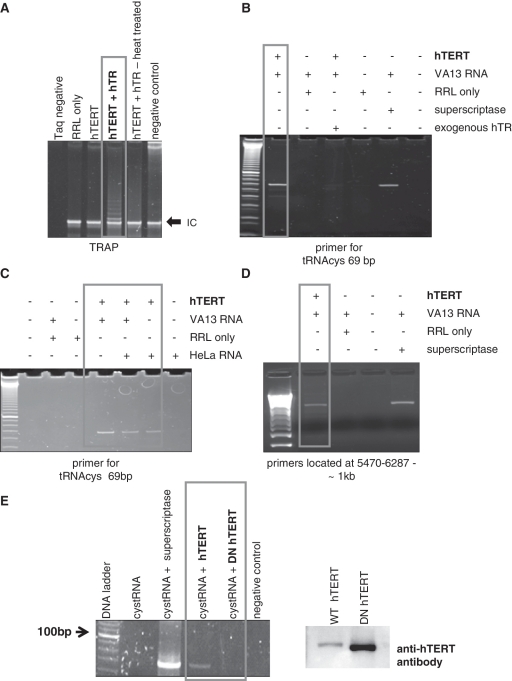
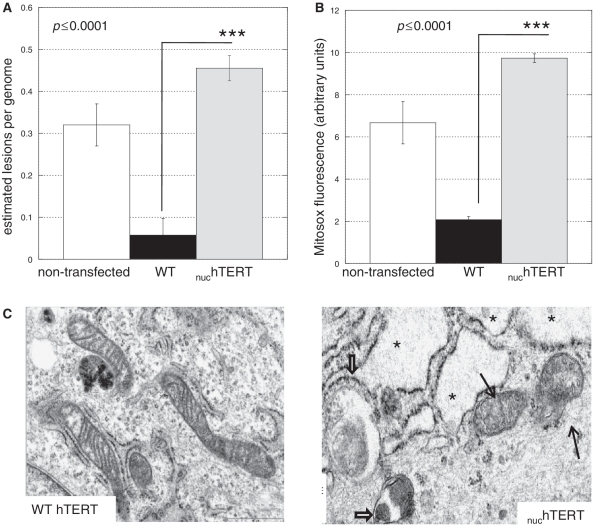
Human telomerase reverse transcriptase (hTERT) is localized to mitochondria, as well as the nucleus, but details about its biology and function in the organelle remain largely unknown. Here we show, using multiple approaches, that mammalian TERT is mitochondrial, co-purifying with mitochondrial nucleoids and tRNAs. We demonstrate the canonical nuclear RNA [human telomerase RNA (hTR)] is not present in human mitochondria and not required for the mitochondrial effects of telomerase, which nevertheless rely on reverse transcriptase (RT) activity. Using RNA immunoprecipitations from whole cell and in organello, we show that hTERT binds various mitochondrial RNAs, suggesting that RT activity in the organelle is reconstituted with mitochondrial RNAs. In support of this conclusion, TERT drives first strand cDNA synthesis in vitro in the absence of hTR. Finally, we demonstrate that absence of hTERT specifically in mitochondria with maintenance of its nuclear function negatively impacts the organelle. Our data indicate that mitochondrial hTERT works as a hTR-independent reverse transcriptase, and highlight that nuclear and mitochondrial telomerases have different cellular functions. The implications of these findings to both the mitochondrial and telomerase fields are discussed.
Figures
References
-
- Santos JH, Meyer JN, Skorvaga M, Annab LA, Van Houten B. Mitochondrial hTERT exacerbates free radical-mediated mtDNA damage. Aging Cell. 2004;6:399–411. - PubMed
-
- Santos JH, Meyer JN, Van Houten B. Mitochondrial localization of hTERT as a determinant for hydrogen peroxide-induced mtDNA damage and apoptosis. Human Mol. Gen. 2006;15:1757–1768. - PubMed
-
- Ahmed S, Passos JF, Birket MJ, Beckmann T, Brings S, Peters H, Birch-Machin MA, von Zglinicki T, Saretzki G. Telomerase does not counteract telomere shortening but protects mitochondrial function under oxidative stress. J. Cell. Sci. 2008;121:1046–1053. - PubMed
-
- Haendeler J, Dröse S, Büchner N, Jakob S, Altschmied J, Goy C, Spyridopoulos I, Zeiher AM, Brandt U, Dimmeler S. Mitochondrial telomerase reverse transcriptase binds to and protects mitochondrial DNA and function from damage. Arterioscler. Thromb. Vasc. Biol. 2009;6:929–935. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
