

Review
doi: 10.1101/gad.17420111.
AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function
Affiliations
- PMID: 21937710
- PMCID: PMC3185962
- DOI: 10.1101/gad.17420111
Item in Clipboard
Review
AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function
Genes Dev.
.
Abstract
AMP-activated protein kinase (AMPK) is a sensor of energy status that maintains cellular energy homeostasis. It arose very early during eukaryotic evolution, and its ancestral role may have been in the response to starvation. Recent work shows that the kinase is activated by increases not only in AMP, but also in ADP. Although best known for its effects on metabolism, AMPK has many other functions, including regulation of mitochondrial biogenesis and disposal, autophagy, cell polarity, and cell growth and proliferation. Both tumor cells and viruses establish mechanisms to down-regulate AMPK, allowing them to escape its restraining influences on growth.
Figures
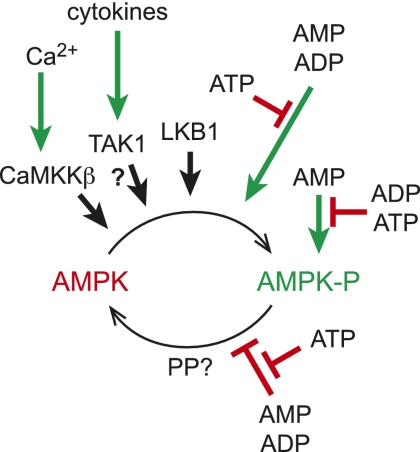
Regulation of AMPK by phosphorylation and adenine nucleotides. AMPK is activated >200-fold by phosphorylation at Thr 172, catalyzed by three upstream kinases: (1) LKB1, which appears to be constitutively active; (2) TAK1, which is activated by cytokines (as the physiological significance of this remains uncertain, it is shown with a question mark); and (3) CaMKKβ, which is activated by a rise in cytosol Ca2+. Phosphorylation is stimulated and dephosphorylation is inhibited by conformational changes triggered by binding of AMP or ADP to the γ subunit of AMPK, both effects being antagonized by ATP. AMP binding also causes a further 10-fold allosteric activation; this effect is antagonized by both ATP and ADP. The identity of the protein phosphatase that dephosphorylates Thr 172 (PP?) remains unclear.

Alignment of consensus site for recognition of substrates by AMPK (Scott et al. 2002) with the sequences (from humans) of some established physiological substrates. In the consensus sequence, hydrophobic residues are represented by Φ and basic residues are represented by β. AMPK phosphorylates serine residues (threonine also allowed) in the context of hydrophobic residues (in bold, red) at P-5 (usually M or L, but I, V, or F also allowed) and P+4 (usually L, but I, M, F, V, Q, or N allowed), and basic residues (in bold, blue; usually R, but K or H allowed) at P-4, P-3, or both. Another basic residue at P-6 is also a positive determinant, although not essential (Scott et al. 2002). Some substrates (e.g., ACC1) also have hydrophobic side chains at regular spacings of three to four residues (bold, red) running in an N-terminal direction from P-5, which form an amphipathic helix. This is clearly not essential, since the sequence of muscle glycogen synthase starts at P-7. Note that one substrate (PFKFB2) still has a βΦβ motif, but this is at P-5/P-4/P-3 rather than the more common P-6/P-5/P-4.
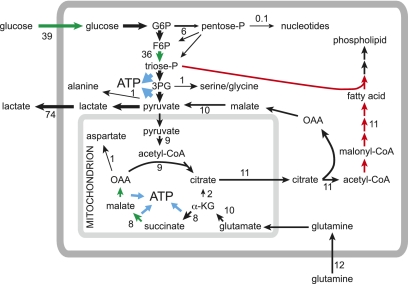
Metabolism in rapidly proliferating cells as revealed by metabolic flux analysis in the lung adenocarcinoma cell line A549, grown in medium containing 25 mM glucose and 4 mM glutamine (note that A549 cells do not express LKB1, so AMPK activity would be very low) (Metallo et al. 2009). Numbers refer to the estimated fluxes in nanomole per minute per milligram protein. Note that the flux out of the TCA cycle into fatty acid synthesis (presumably required for synthesis of new membrane phospholipid) is greater than the flux entering the TCA cycle from pyruvate. This deficit is made up by anaplerotic flux from glutamine, with the flow from α-ketoglutarate (αKG) to citrate operating in the reverse of the usual direction of the TCA cycle. Note also the very large flux through glucose uptake, glycolysis, and lactate production. Processes currently known to be down-regulated by AMPK activation are shown in red, and processes known to be up-regulated by AMPK are in green. In general, AMPK switches off anabolic (ATP-consuming) processes and switches on catabolic (ATP-producing) processes.
References
-
- Amodeo GA, Rudolph MJ, Tong L 2007. Crystal structure of the heterotrimer core of Saccharomyces cerevisiae AMPK homologue SNF1. Nature 449: 492–495 - PubMed
-
- Baas AF, Kuipers J, van der Wel NN, Batlle E, Koerten HK, Peters PJ, Clevers HC 2004. Complete polarization of single intestinal epithelial cells upon activation of LKB1 by STRAD. Cell 116: 457–466 - PubMed
-
- Baena-Gonzalez E, Rolland F, Thevelein JM, Sheen J 2007. A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942 - PubMed
-
- Barnes K, Ingram JC, Porras OH, Barros LF, Hudson ER, Fryer LG, Foufelle F, Carling D, Hardie DG, Baldwin SA 2002. Activation of GLUT1 by metabolic and osmotic stress: potential involvement of AMP-activated protein kinase (AMPK). J Cell Sci 115: 2433–2442 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources