

Overview of the matrisome--an inventory of extracellular matrix constituents and functions
- PMID: 21937732
- PMCID: PMC3249625
- DOI: 10.1101/cshperspect.a004903
Overview of the matrisome--an inventory of extracellular matrix constituents and functions
Abstract
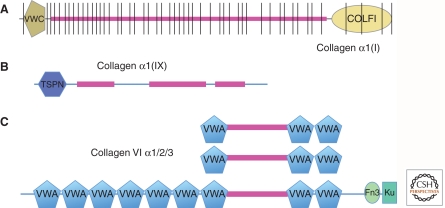
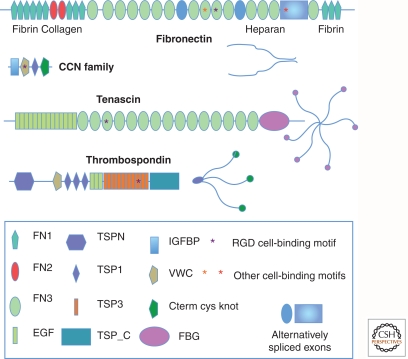
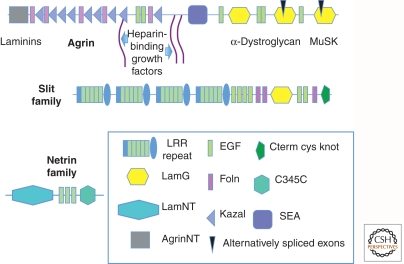
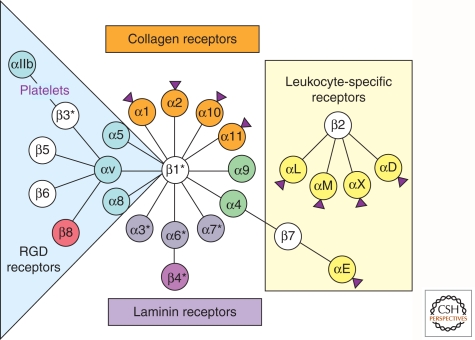
Completion of genome sequences for many organisms allows a reasonably complete definition of the complement of extracellular matrix (ECM) proteins. In mammals this "core matrisome" comprises ∼300 proteins. In addition there are large numbers of ECM-modifying enzymes, ECM-binding growth factors, and other ECM-associated proteins. These different categories of ECM and ECM-associated proteins cooperate to assemble and remodel extracellular matrices and bind to cells through ECM receptors. Together with receptors for ECM-bound growth factors, they provide multiple inputs into cells to control survival, proliferation, differentiation, shape, polarity, and motility of cells. The evolution of ECM proteins was key in the transition to multicellularity, the arrangement of cells into tissue layers, and the elaboration of novel structures during vertebrate evolution. This key role of ECM is reflected in the diversity of ECM proteins and the modular domain structures of ECM proteins both allow their multiple interactions and, during evolution, development of novel protein architectures.
Figures
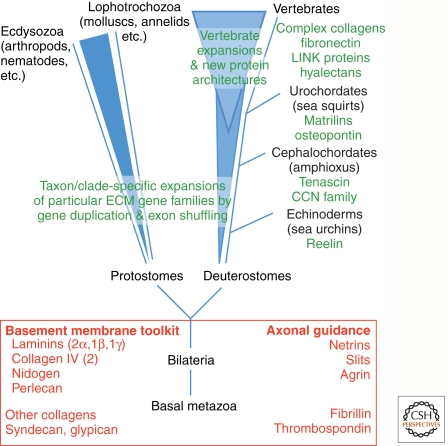
References
-
- Adams J, Engel J 2007. Bioinformatic analysis of adhesion proteins. In Methods in molecular biology, pp. 147–172 Humana Press, New York - PubMed
-
- Aszódi A, Legate KR, Nakchbandi I, Fässler R 2006. What mouse mutants teach us about extracellular matrix function. Annu Rev Cell Dev Biol 22: 591–621 - PubMed
-
- Aumailley M, Bruckner-Tuderman L, Carter WG, Deutzmann R, Edgar D, Ekblom P, Engel J, Engvall E, Hohenester E, Jones JCR, et al. 2005. A simplified laminin nomenclature. Matrix Biol 24: 326–332 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources