

The magnitude of the T cell response to a clinically significant dose of influenza virus is regulated by TRAIL
- PMID: 21940678
- PMCID: PMC3197947
- DOI: 10.4049/jimmunol.1002241
The magnitude of the T cell response to a clinically significant dose of influenza virus is regulated by TRAIL
Abstract
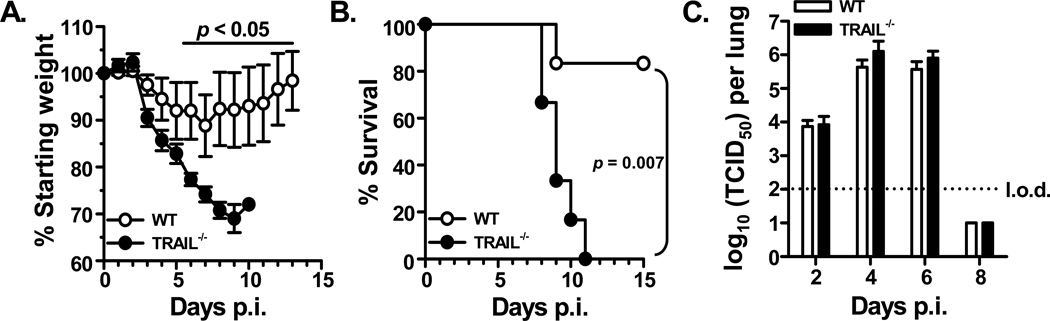
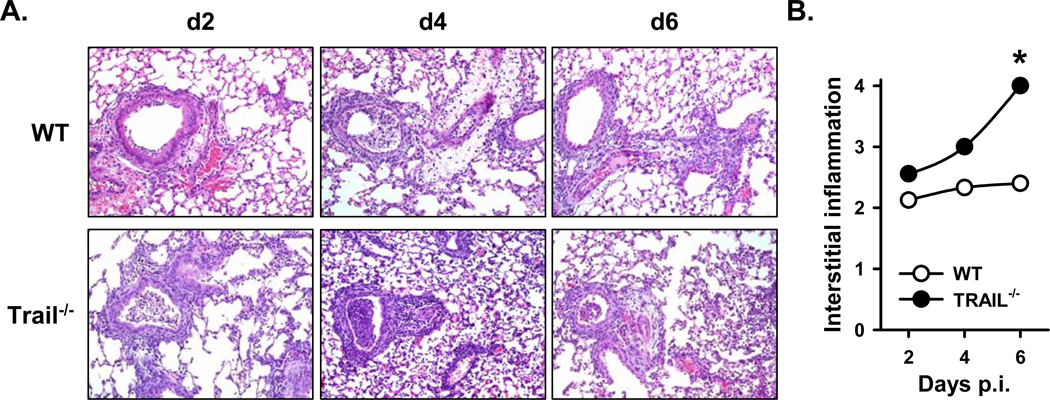
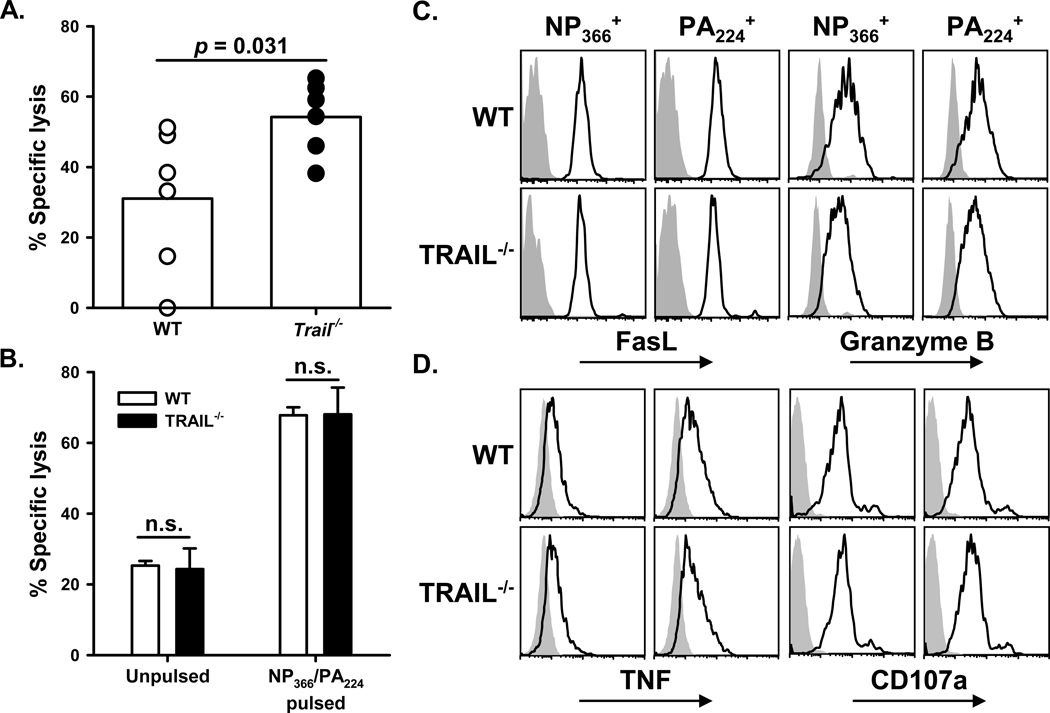
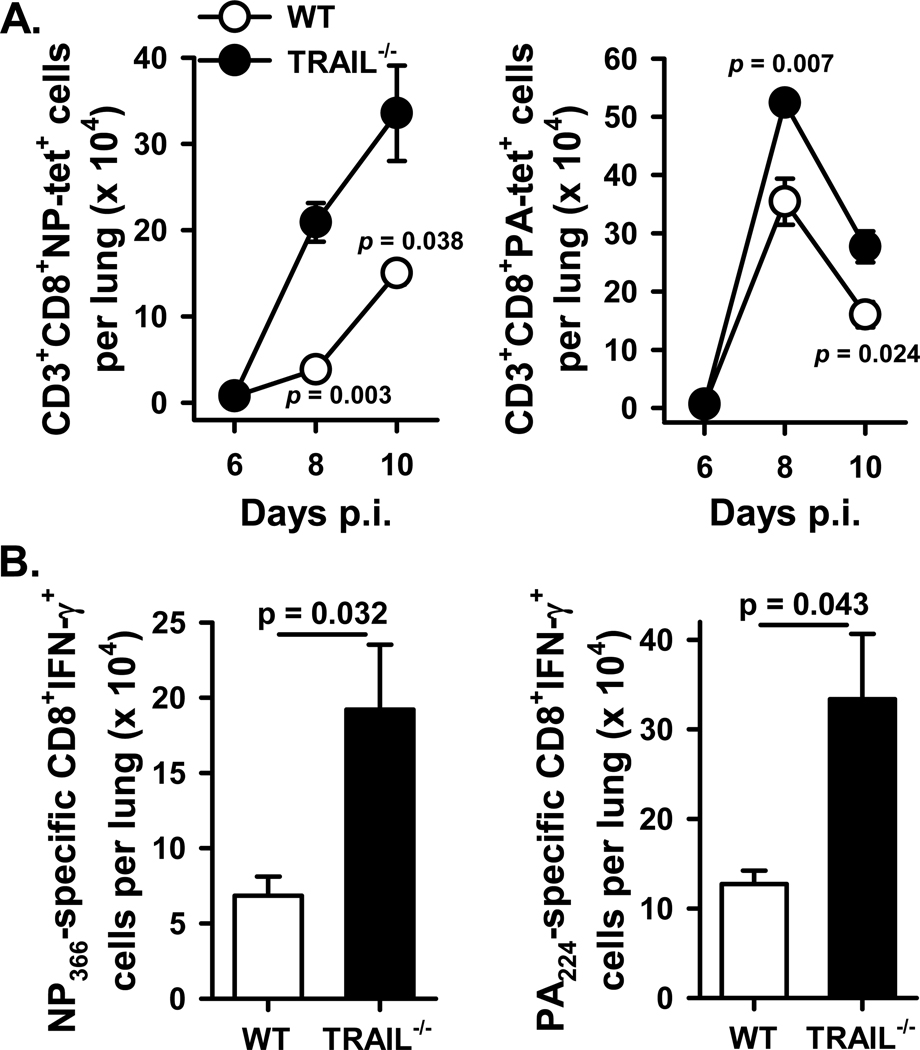
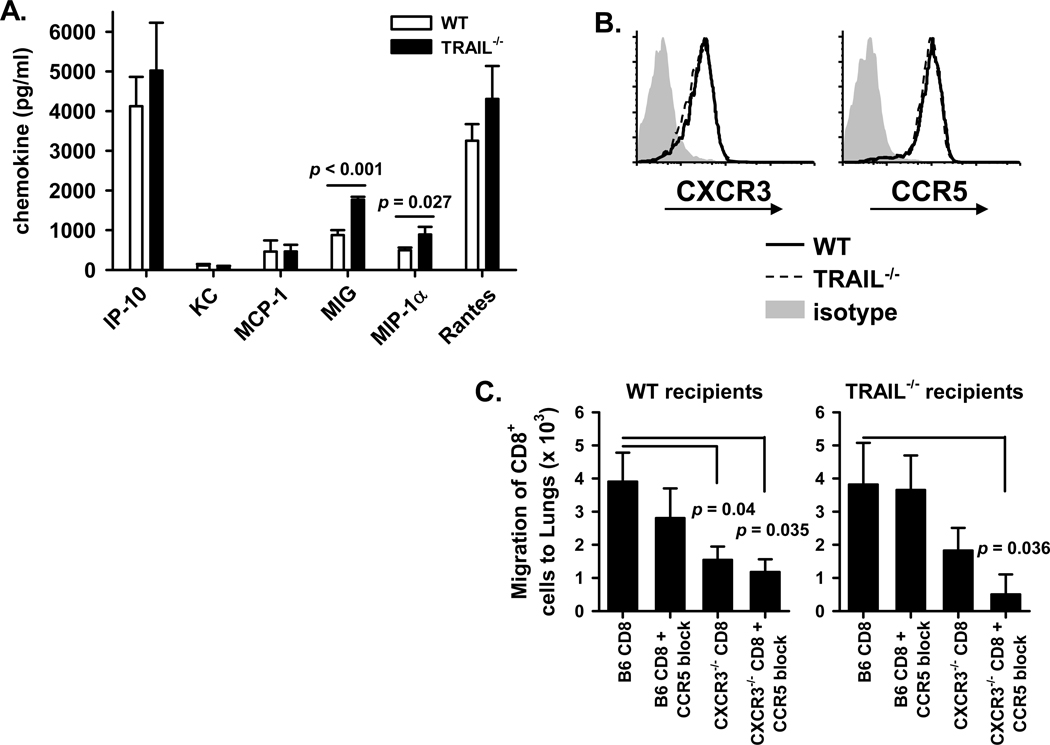
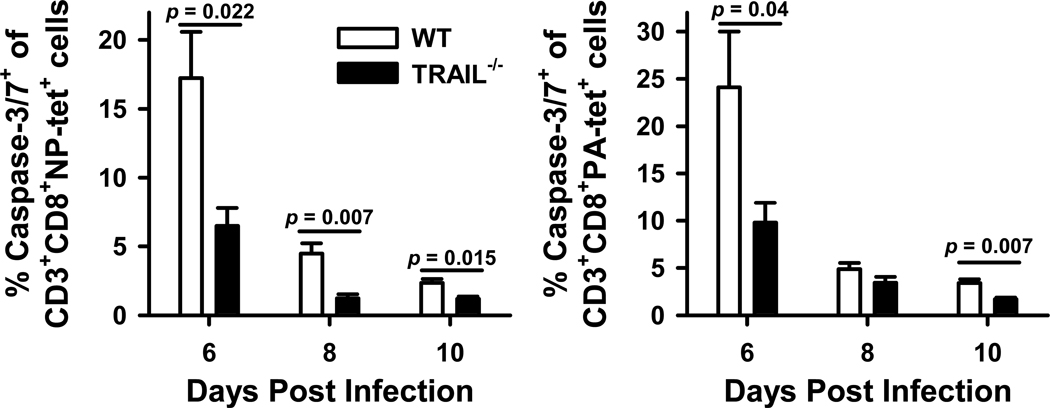
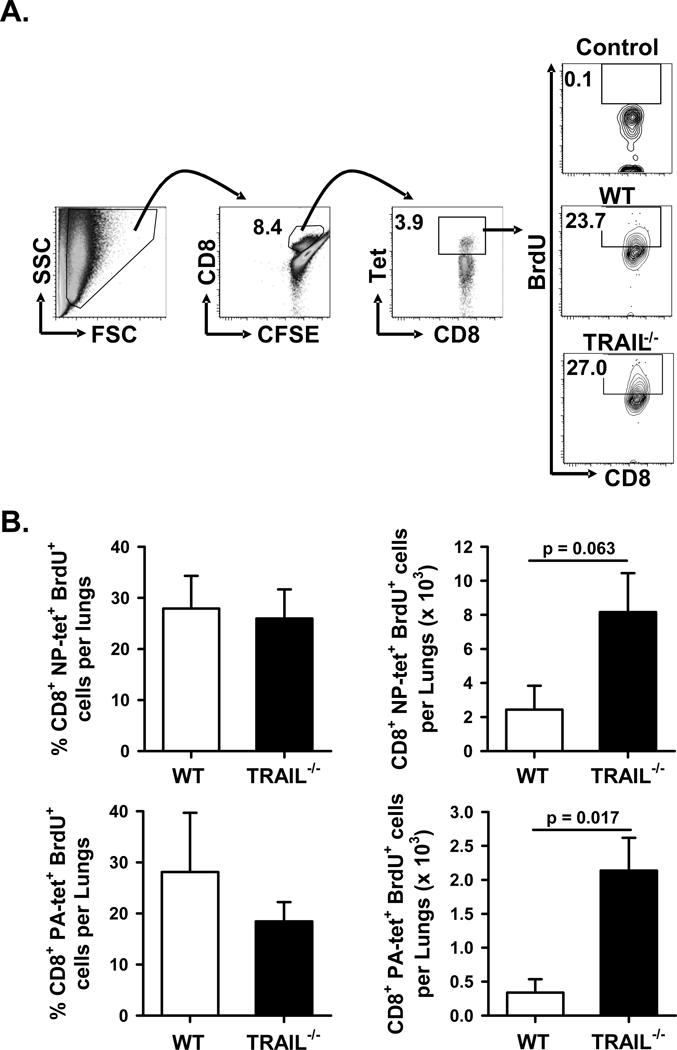
An immune response of appropriate magnitude should be robust enough to control pathogen spread but not simultaneously lead to immunopathology. Primary infection with influenza A virus (IAV) results in a localized pulmonary infection and inflammation and elicits an IAV-specific CD8 T cell immune response necessary for viral clearance. Clearance of IAV-infected cells, and recovery from infection, is mediated by perforin/granzyme B- and Fas/FasL-mediated mechanisms. We recently reported that TRAIL is another means by which IAV-specific CD8 T cells can kill IAV-infected cells. The current study examined the role of TRAIL in the pulmonary CD8 T cell response to a clinically significant IAV [A/PR/8/34 (PR8; H1N1)] infection (i.e., leads to observable, but limited, morbidity and mortality in wild-type [WT] mice). Compared with WT mice, IAV-infected Trail(-/-) mice experienced increased morbidity and mortality despite similar rates of viral clearance from the lungs. The increased morbidity and mortality in Trail(-/-) mice correlated with increased pulmonary pathology and inflammatory chemokine production. Analysis of lung-infiltrating lymphocytes revealed increased numbers of IAV-specific CD8 T cells in infected Trail(-/-) mice, which correlated with increased pulmonary cytotoxic activity and increased pulmonary expression of MIG and MIP-1α. In addition, there was decreased apoptosis and increased proliferation of IAV-specific CD8 T cells in the lungs of Trail(-/-) mice compared with WT mice. Together, these data suggest that TRAIL regulates the magnitude of the IAV-specific CD8 T cell response during a clinically significant IAV infection to decrease the chance for infection-induced immunopathology.
Figures
References
-
- Epstein SL, Lo CY, Misplon JA, Bennink JR. Mechanism of protective immunity against influenza virus infection in mice without antibodies. J. Immunol. 1998;160:322–327. - PubMed
-
- Swain SL, Dutton RW, Woodland DL. T cell responses to influenza virus infection: effector and memory cells. Viral Immunol. 2004;17:197–209. - PubMed
-
- Lawrence CW, Braciale TJ. Activation, differentiation, and migration of naive virus-specific CD8+ T cells during pulmonary influenza virus infection. J. Immunol. 2004;173:1209–1218. - PubMed
-
- Lawrence CW, Ream RM, Braciale TJ. Frequency, specificity, and sites of expansion of CD8+ T cells during primary pulmonary influenza virus infection. J. Immunol. 2005;174:5332–5340. - PubMed
-
- Legge KL, Braciale TJ. Lymph node dendritic cells control CD8+ T cell responses through regulated FasL expression. Immunity. 2005;23:649–659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
