

Functional characterization of barley betaglucanless mutants demonstrates a unique role for CslF6 in (1,3;1,4)-β-D-glucan biosynthesis
- PMID: 21940720
- PMCID: PMC3245474
- DOI: 10.1093/jxb/err285
Functional characterization of barley betaglucanless mutants demonstrates a unique role for CslF6 in (1,3;1,4)-β-D-glucan biosynthesis
Abstract
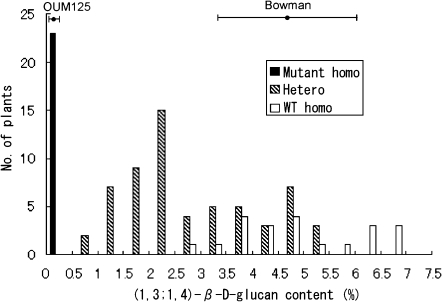
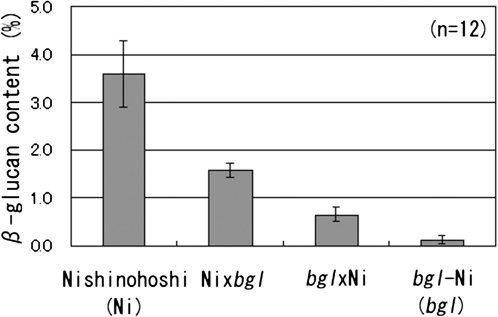
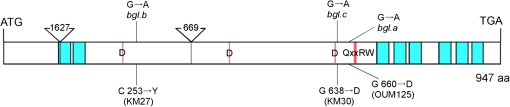
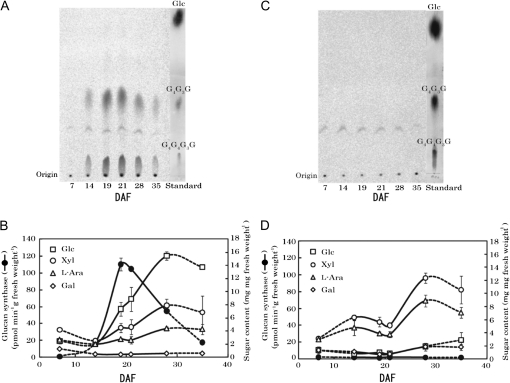
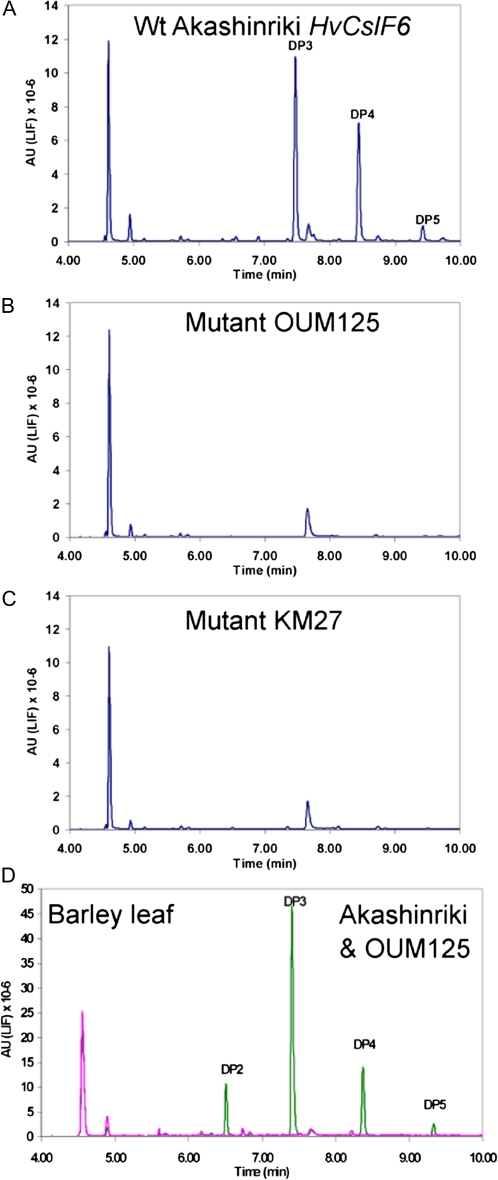
(1,3;1,4)-β-D-glucans (mixed-linkage glucans) are found in tissues of members of the Poaceae (grasses), and are particularly high in barley (Hordeum vulgare) grains. The present study describes the isolation of three independent (1,3;1,4)-β-D-glucanless (betaglucanless; bgl) mutants of barley which completely lack (1,3;1,4)-β-D-glucan in all the tissues tested. The bgl phenotype cosegregates with the cellulose synthase like HvCslF6 gene on chromosome arm 7HL. Each of the bgl mutants has a single nucleotide substitution in the coding region of the HvCslF6 gene resulting in a change of a highly conserved amino acid residue of the HvCslF6 protein. Microsomal membranes isolated from developing endosperm of the bgl mutants lack detectable (1,3;1,4)-β-D-glucan synthase activity indicating that the HvCslF6 protein is inactive. This was confirmed by transient expression of the HvCslF6 cDNAs in Nicotiana benthamiana leaves. The wild-type HvCslF6 gene directed the synthesis of high levels of (1,3;1,4)-β-D-glucans, whereas the mutant HvCslF6 proteins completely lack the ability to synthesize (1,3;1,4)-β-D-glucans. The fine structure of the (1,3;1,4)-β-D-glucan produced in the tobacco leaf was also very different from that found in cereals having an extremely low DP3/DP4 ratio. These results demonstrate that, among the seven CslF and one CslH genes present in the barley genome, HvCslF6 has a unique role and is the key determinant controlling the biosynthesis of (1,3;1,4)-β-D-glucans. Natural allelic variation in the HvCslF6 gene was found predominantly within introns among 29 barley accessions studied. Genetic manipulation of the HvCslF6 gene could enable control of (1,3;1,4)-β-D-glucans in accordance with the purposes of use.
Figures

References
-
- Bar-Peled M, O'Neill MA. Plant nucleotide sugar formation, interconversion, and salvage by sugar recycling. Annual Review of Plant Biology. 2011;62:127–155. - PubMed
-
- Bouveng HS, Lindberg B. Hydrolysis of methylated polysaccharides. Methods in Carbohydrate Chemistry. 1965;5:269–276.
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Analytical Biochemistry. 1976;72:248–254. - PubMed
-
- Brennan CS, Cleary LJ. The potential use of (1→3,1→4)-β-D-glucans as functional food ingredients. Journal of Cereal Science. 2005;42:1–13.
-
- Buckeridge MS, Rayon C, Urbanowicz B, Tine MAS, Carpita NC. Mixed linkage (1→3),(1→4)-β-D-glucans of grasses. Cereal Chemistry. 2004;81:115–127.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
