

Cofilin cooperates with fascin to disassemble filopodial actin filaments
- PMID: 21940796
- PMCID: PMC4074248
- DOI: 10.1242/jcs.086934
Cofilin cooperates with fascin to disassemble filopodial actin filaments
Abstract
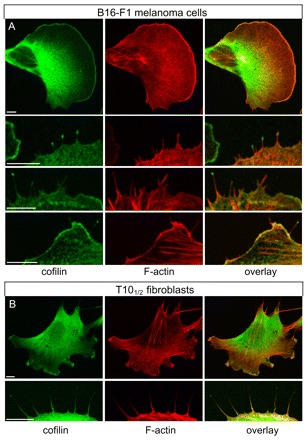
Cells use a large repertoire of proteins to remodel the actin cytoskeleton. Depending on the proteins involved, F-actin is organized in specialized protrusions such as lamellipodia or filopodia, which serve diverse functions in cell migration and sensing. Although factors responsible for directed filament assembly in filopodia have been extensively characterized, the mechanisms of filament disassembly in these structures are mostly unknown. We investigated how the actin-depolymerizing factor cofilin-1 affects the dynamics of fascincrosslinked actin filaments in vitro and in live cells. By multicolor total internal reflection fluorescence microscopy and fluorimetric assays, we found that cofilin-mediated severing is enhanced in fascin-crosslinked bundles compared with isolated filaments, and that fascin and cofilin act synergistically in filament severing. Immunolabeling experiments demonstrated for the first time that besides its known localization in lamellipodia and membrane ruffles, endogenous cofilin can also accumulate in the tips and shafts of filopodia. Live-cell imaging of fluorescently tagged proteins revealed that cofilin is specifically targeted to filopodia upon stalling of protrusion and during their retraction. Subsequent electron tomography established filopodial actin filament and/or bundle fragmentation to precisely correlate with cofilin accumulation. These results identify a new mechanism of filopodium disassembly involving both fascin and cofilin.
@ 2011. Published by The Company of Biologists Ltd
Figures








References
-
- Auinger S., Small J. V. (2008). Correlated light and electron microscopy of the cytoskeleton. Methods in Cell Biol. 88, 257-272 - PubMed
-
- Andrianantoandro E., Pollard T. D. (2006). Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell 24, 13-23 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
