

Highlights of 10 years of immunology in Nature Reviews Immunology
- PMID: 21941295
- PMCID: PMC3703536
- DOI: 10.1038/nri3063
Highlights of 10 years of immunology in Nature Reviews Immunology
Abstract
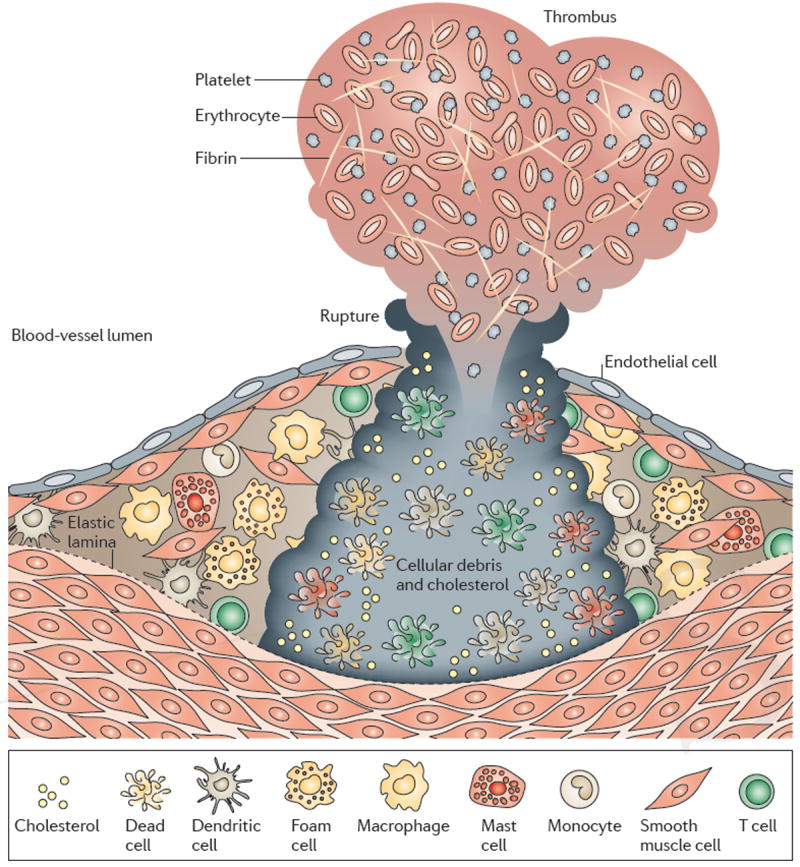
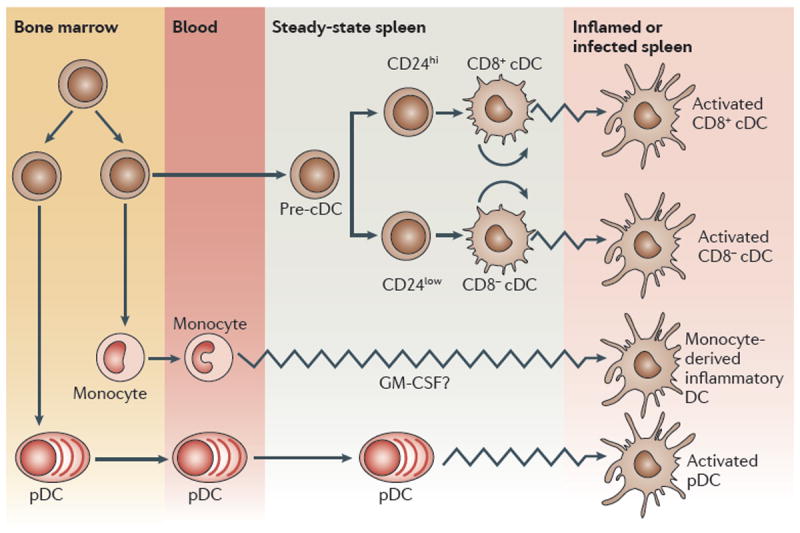
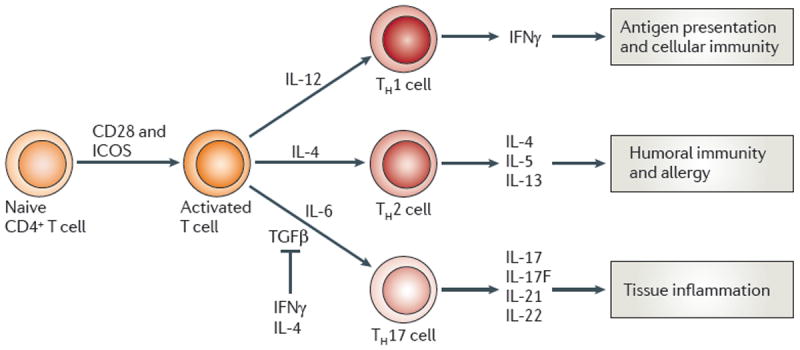
As Nature Reviews Immunology reaches its 10(th) anniversary, the authors of one of the top-cited articles from each year take a trip down memory lane. We've asked them to look back on the state of research at the time their Review was published, to consider why the article has had the impact it has and to discuss the future directions of their field. This Viewpoint article provides an interesting snapshot of some of the fundamental advances in immunology over the past 10 years. Highlights include our improved understanding of Toll-like receptor signalling, and of immune regulation mediated by regulatory T cells, indoleamine 2,3-dioxygenase, myeloid-derived suppressor cells and interleukin-10. The complexities in the development and heterogeneity of macrophages, dendritic cells and T helper cells continue to engage immunologists, as do the immune processes involved in diseases such as atherosclerosis. We look forward to what the next 10 years of immunology research may bring.
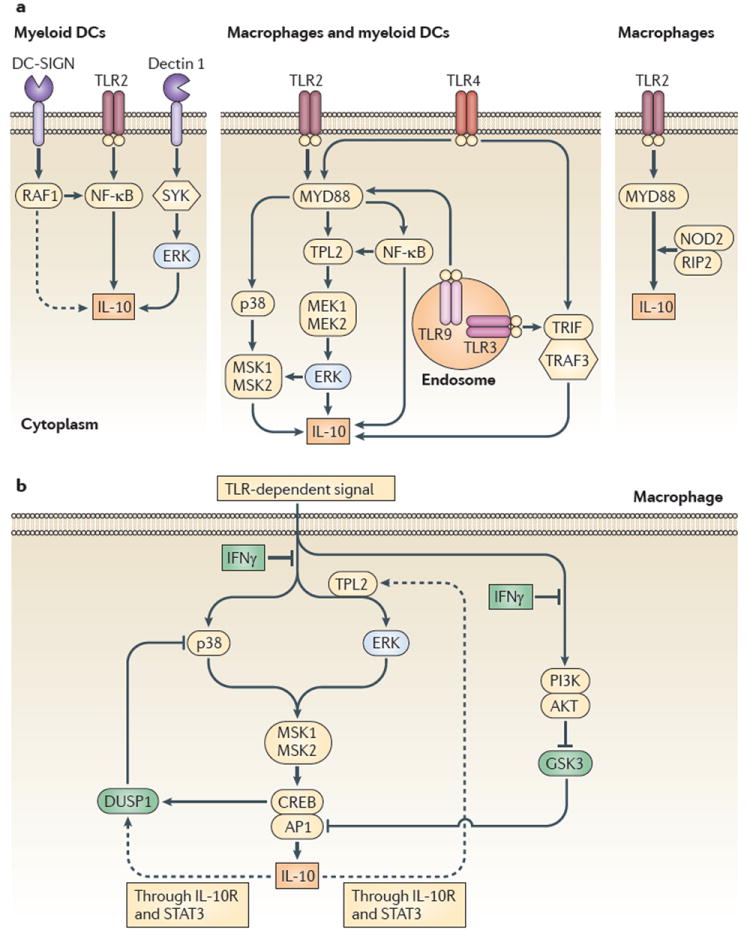
Figures
References
-
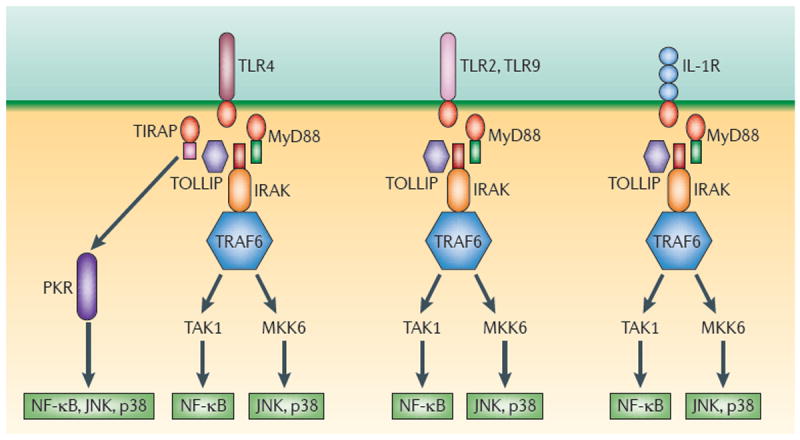
- Medzhitov R. Toll-like receptors and innate immunity. Nature Rev Immunol. 2001;1:135–145. - PubMed
-
- Shevach E. CD4+CD25+ suppressor T cells: more questions than answers. Nature Rev Immunol. 2002;2:389–400. - PubMed
-
- Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor α-chains (CD25) J Immunol. 1995;155:1151–1164. - PubMed
-
- Horri S, Nomura T, Sakaguchi S. Control of T regulatory cell development by the transcription factor Foxp3. Science. 2003;299:1057–1062. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
