

Pathogenesis of aortic dilatation in mucopolysaccharidosis VII mice may involve complement activation
- PMID: 21944884
- PMCID: PMC3283036
- DOI: 10.1016/j.ymgme.2011.08.018
Pathogenesis of aortic dilatation in mucopolysaccharidosis VII mice may involve complement activation
Abstract
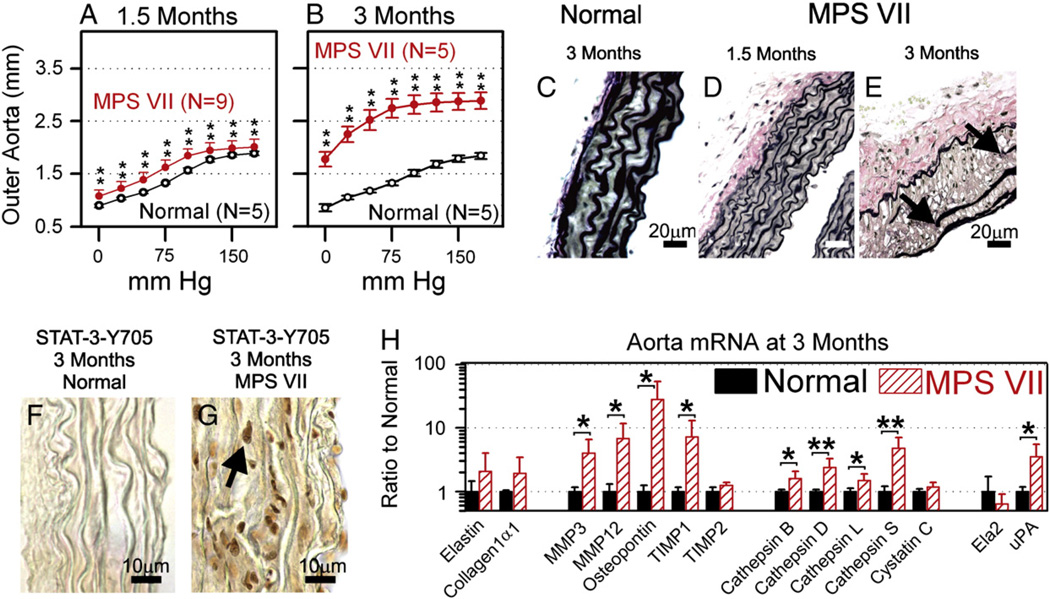
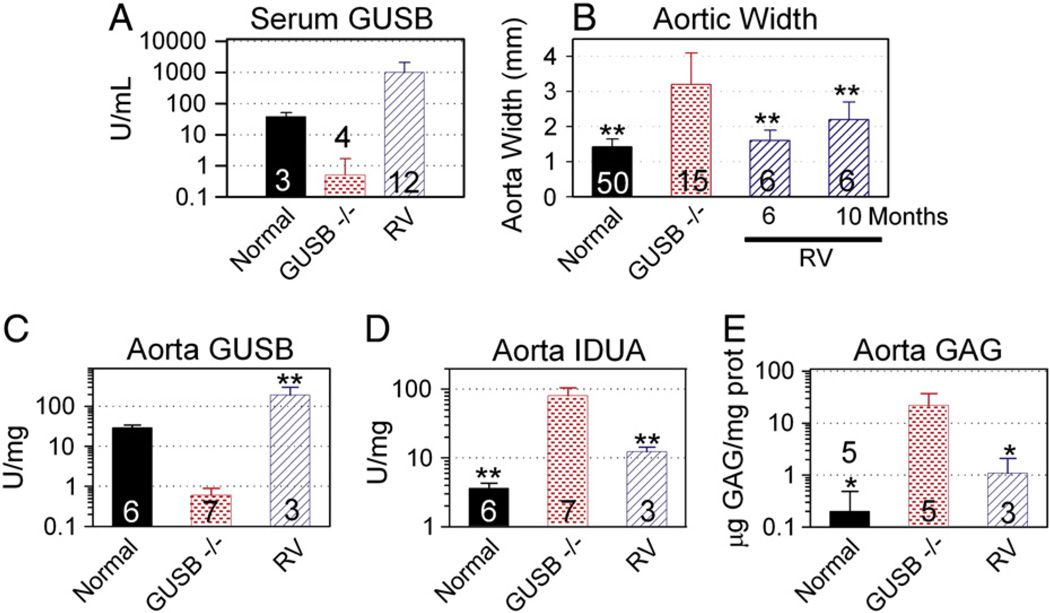
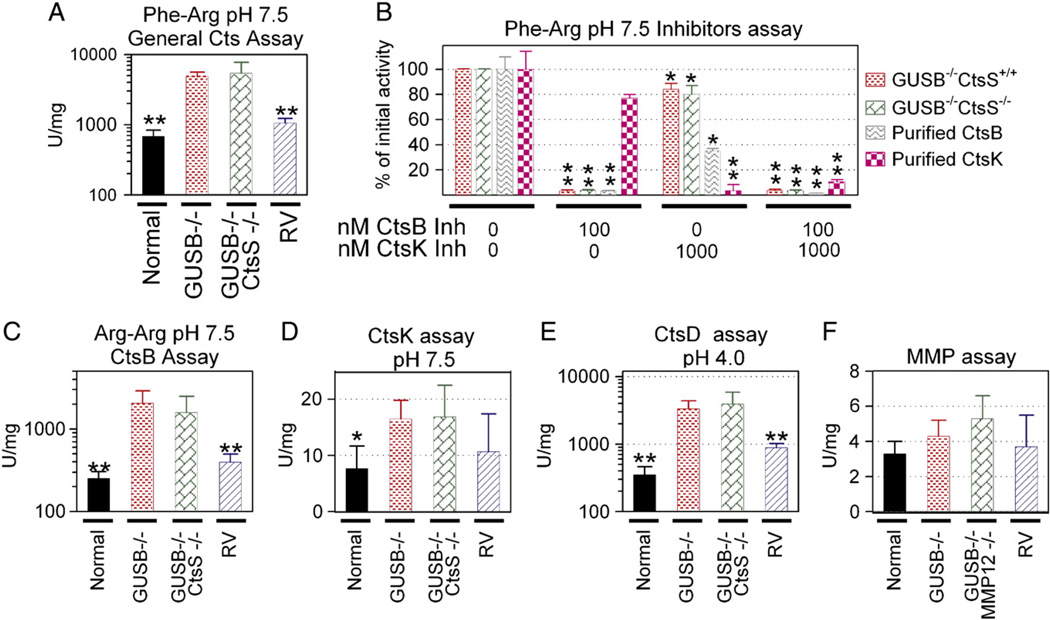
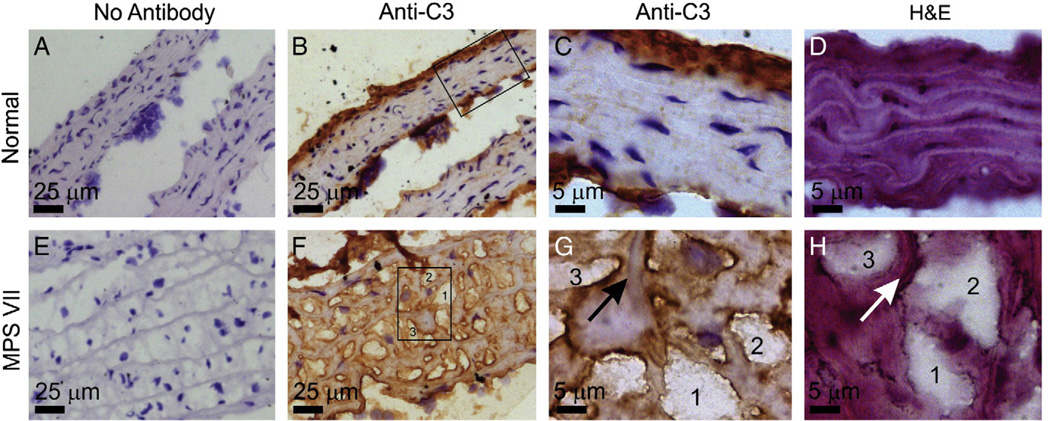
Mucopolysaccharidosis VII (MPS VII) is due to mutations within the gene encoding the lysosomal enzyme β-glucuronidase, and results in the accumulation of glycosaminoglycans. MPS VII causes aortic dilatation and elastin fragmentation, which is associated with upregulation of the elastases cathepsin S (CtsS) and matrix metalloproteinase 12 (MMP12). To test the role of these enzymes, MPS VII mice were crossed with mice deficient in CtsS or MMP12, and the effect upon aortic dilatation was determined. CtsS deficiency did not protect against aortic dilatation in MPS VII mice, but also failed to prevent an upregulation of cathepsin enzyme activity. Further analysis with substrates and inhibitors specific for particular cathepsins suggests that this enzyme activity was due to CtsB, which could contribute to elastin fragmentation. Similarly, MMP12 deficiency and deficiency of both MMP12 and CtsS could not prevent aortic dilatation in MPS VII mice. Microarray and reverse-transcriptase real-time PCR were performed to look for upregulation of other elastases. This demonstrated that mRNA for complement component D was elevated in MPS VII mice, while immunostaining demonstrated high levels of complement component C3 on surfaces within the aortic media. Finally, we demonstrate that neonatal intravenous injection of a retroviral vector encoding β-glucuronidase reduced aortic dilatation. We conclude that neither CtsS nor MMP12 are necessary for elastin fragmentation in MPS VII mouse aorta, and propose that CtsB and/or complement component D may be involved. Complement may be activated by the GAGs that accumulate, and may play a role in signal transduction pathways that upregulate elastases.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


References
-
- Neufeld EF, Muenzer J. The mucopolysaccharidoses. In: Scriver CR, Beaudet AL, Sly WS, Valle D, editors. Metabolic and Molecular Basis of Inherited Disease. New York: McGraw Hill; 2001. pp. 3421–3452.
-
- Ausseil J, Desmaris N, Bigou S, Attali R, Corbineau S, Vitry S, Parent M, Cheillan D, Fuller M, Maire I, Vanier MT, Heard JM. Early neurodegeneration progresses independently of microglial activation by heparan sulfate in the brain of mucopolysaccharidosis IIIB mice. PLoS One. 2008;3:1–11. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
