

Borrelia burgdorferi BBA52 is a potential target for transmission blocking Lyme disease vaccine
- PMID: 21945261
- PMCID: PMC3202674
- DOI: 10.1016/j.vaccine.2011.09.035
Borrelia burgdorferi BBA52 is a potential target for transmission blocking Lyme disease vaccine
Abstract
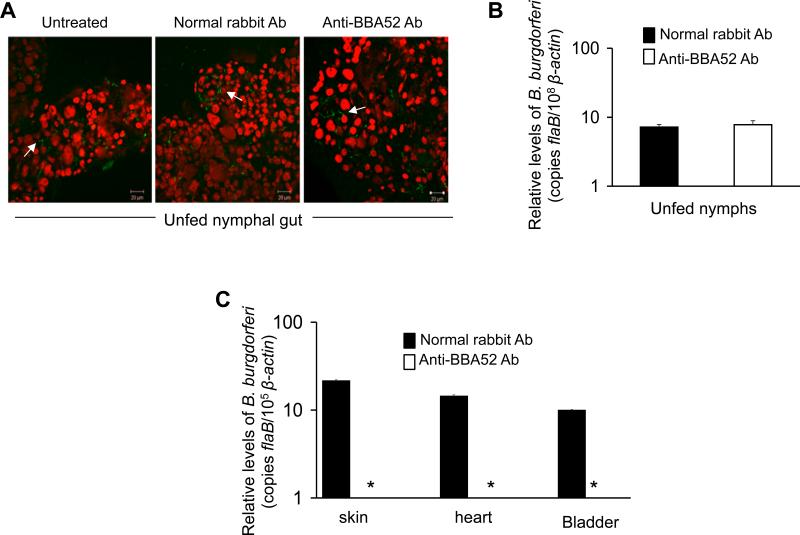
The surface-exposed antigens of Borrelia burgdorferi represent important targets for induction of protective host immune responses. BBA52 is preferentially expressed by B. burgdorferi in the feeding tick, and a targeted deletion of bba52 interferes with vector-host transitions in vivo. In this study, we demonstrate that BBA52 is an outer membrane surface-exposed protein and that disulfide bridges take part in the homo-oligomeric assembly of native protein. BBA52 antibodies lack detectable borreliacidal activities in vitro. However, active immunization studies demonstrated that BBA52 vaccinated mice were significantly less susceptible to subsequent tick-borne challenge infection. Similarly, passive transfer of BBA52 antibodies in ticks completely blocked B. burgdorferi transmission from feeding ticks to naïve mice. Taken together, these studies highlight the role of BBA52 in spirochete dissemination from ticks to mice and demonstrate the potential of BBA52 antibody-mediated strategy to complement the ongoing efforts to develop vaccines for blocking the transmission of B. burgdorferi.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
