

Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus
- PMID: 21946417
- PMCID: PMC3320042
- DOI: 10.1031/ni.2131
Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus
Abstract
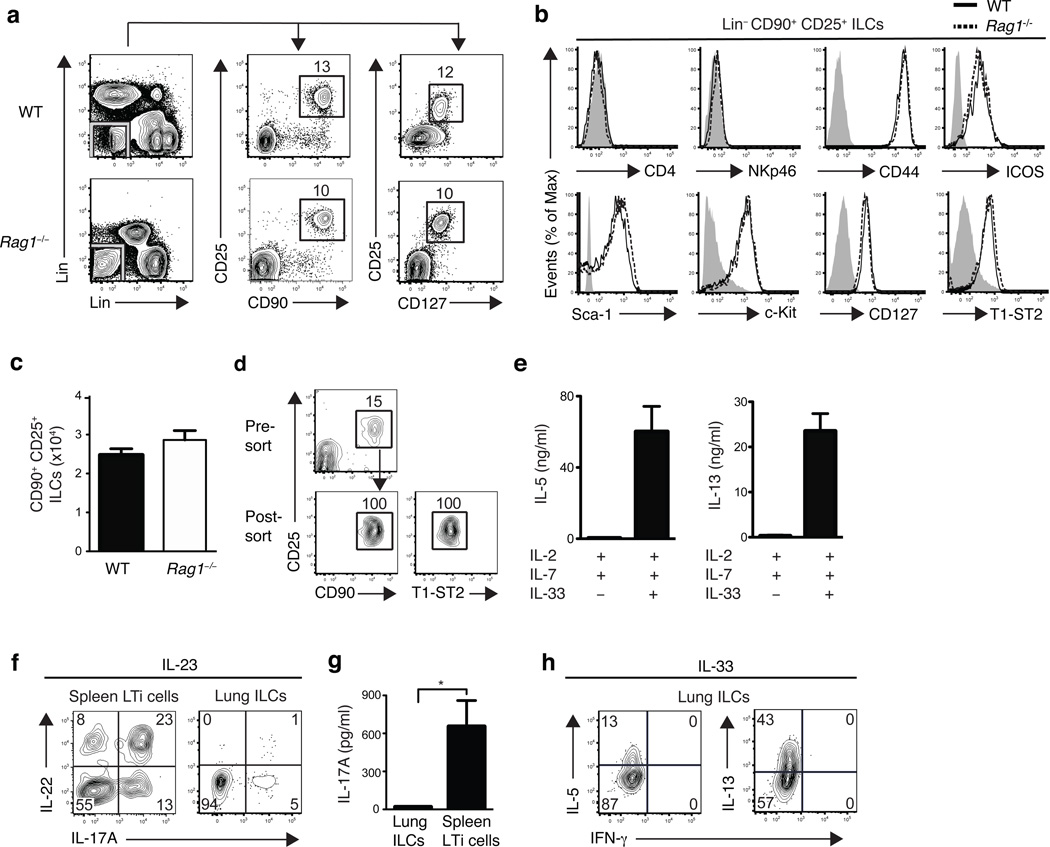
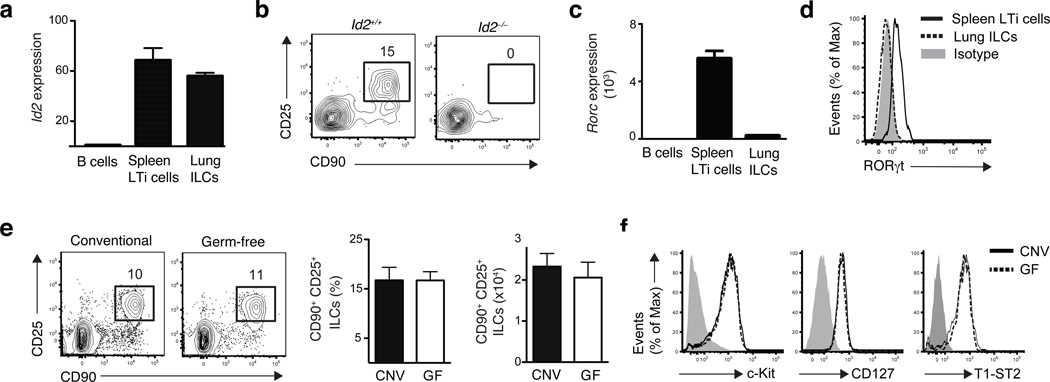
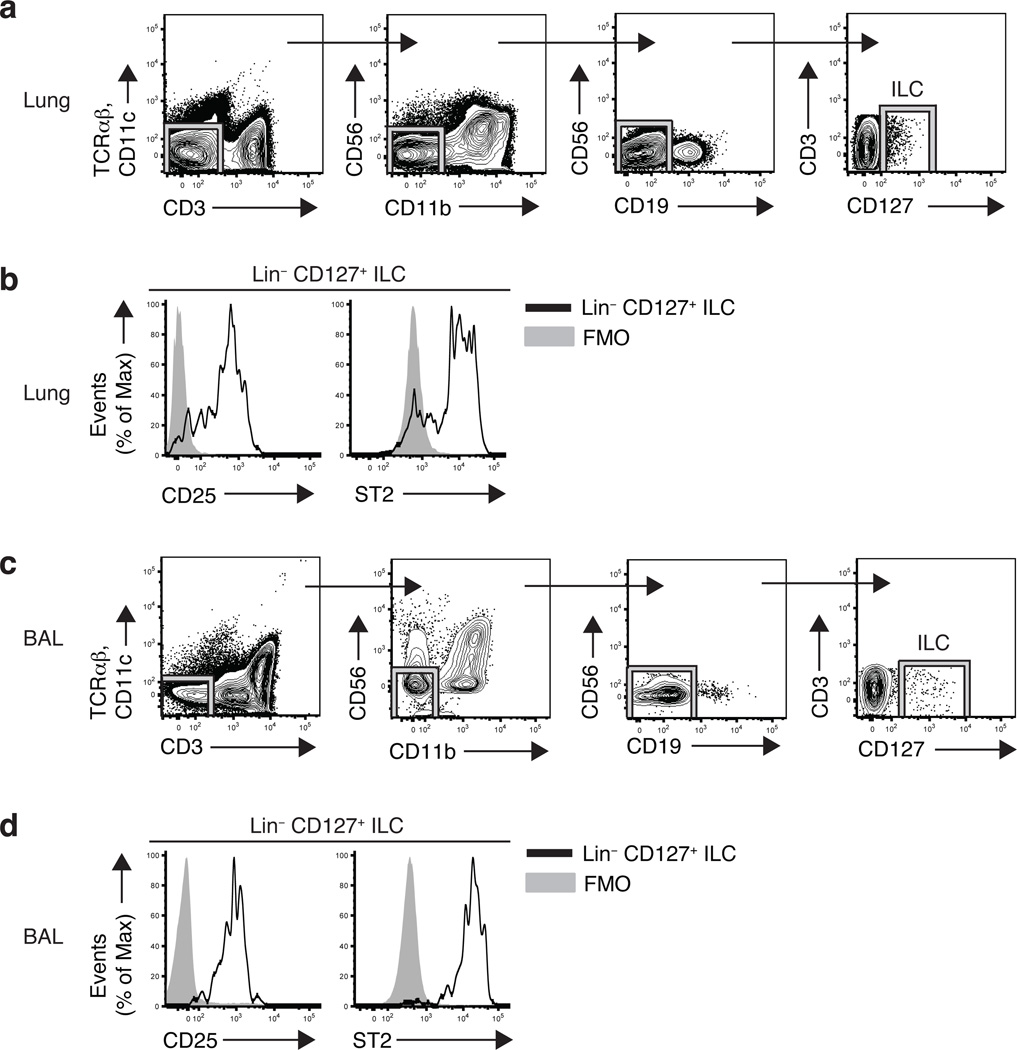
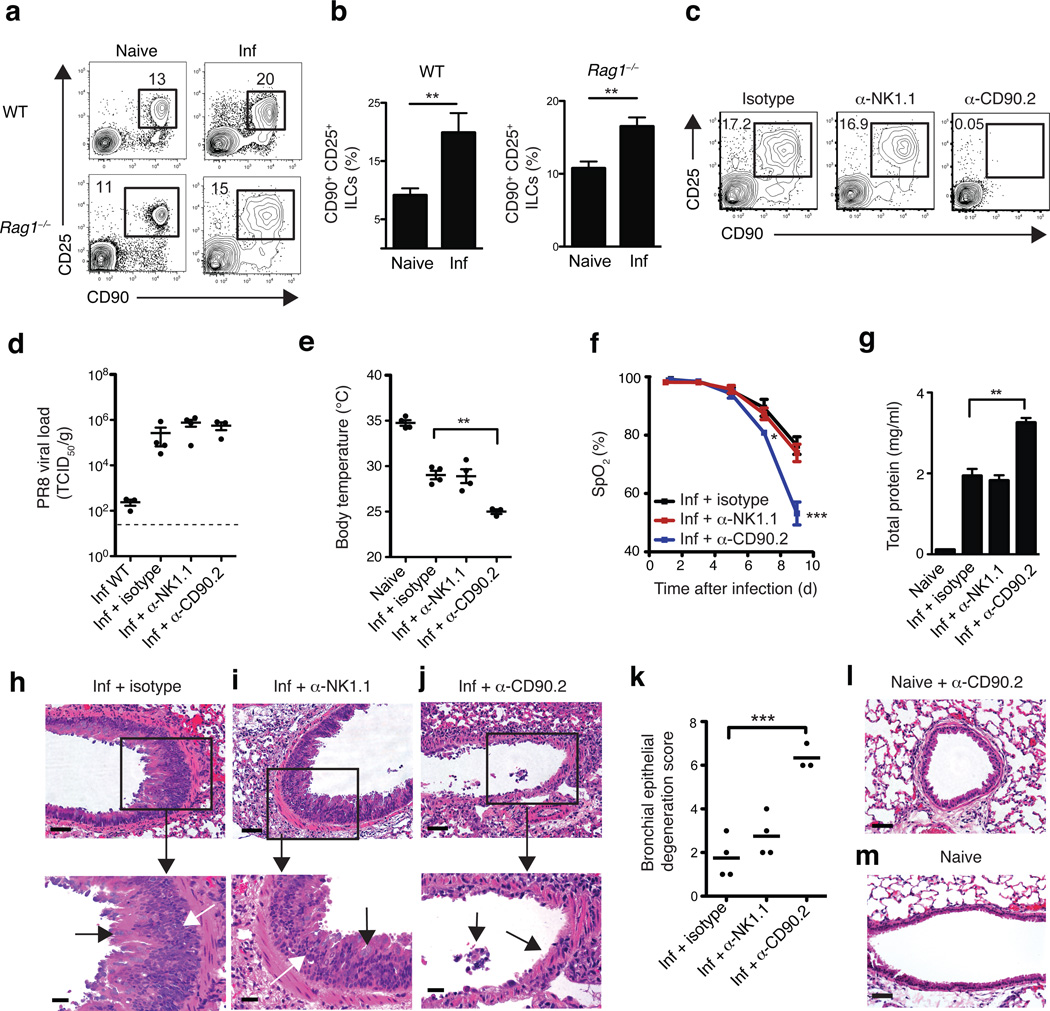
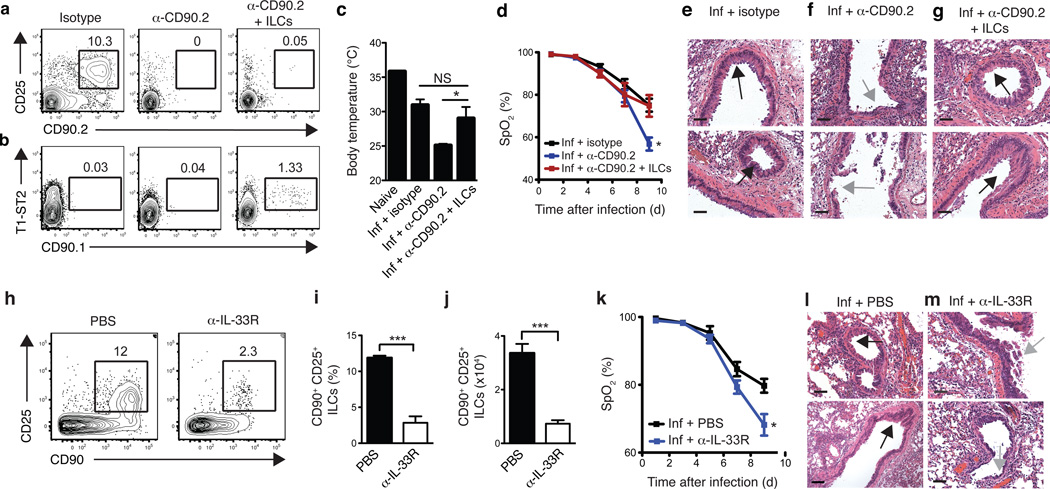
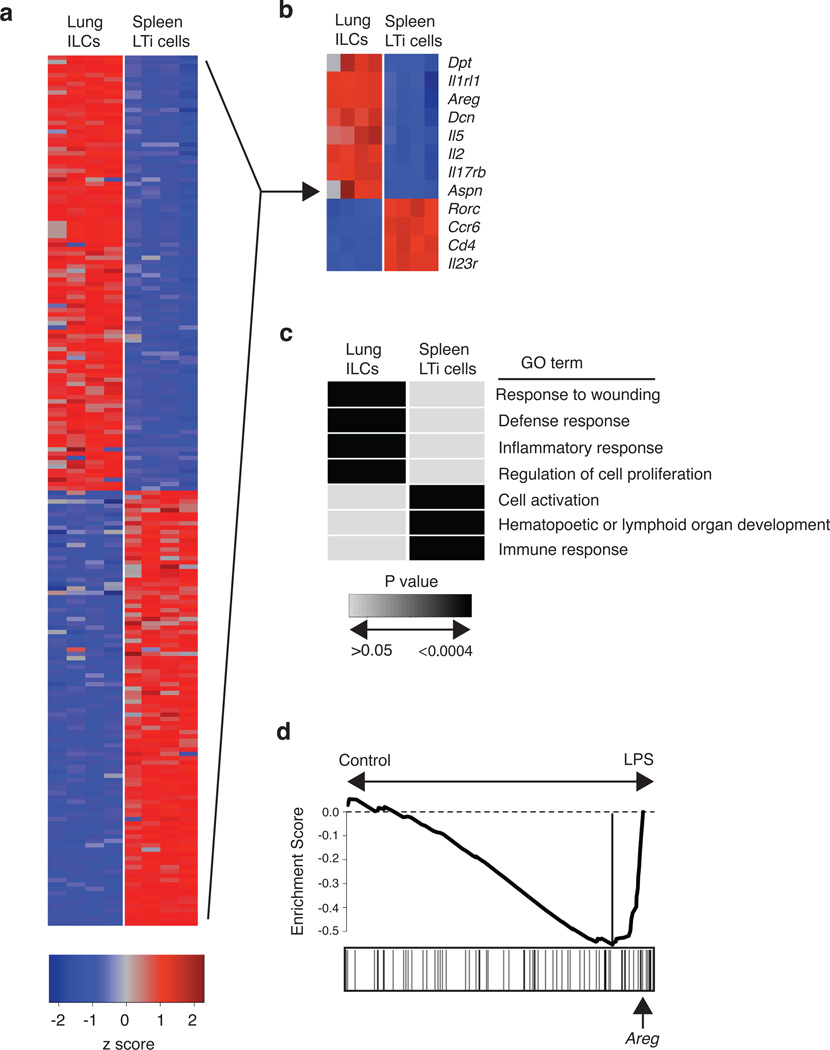
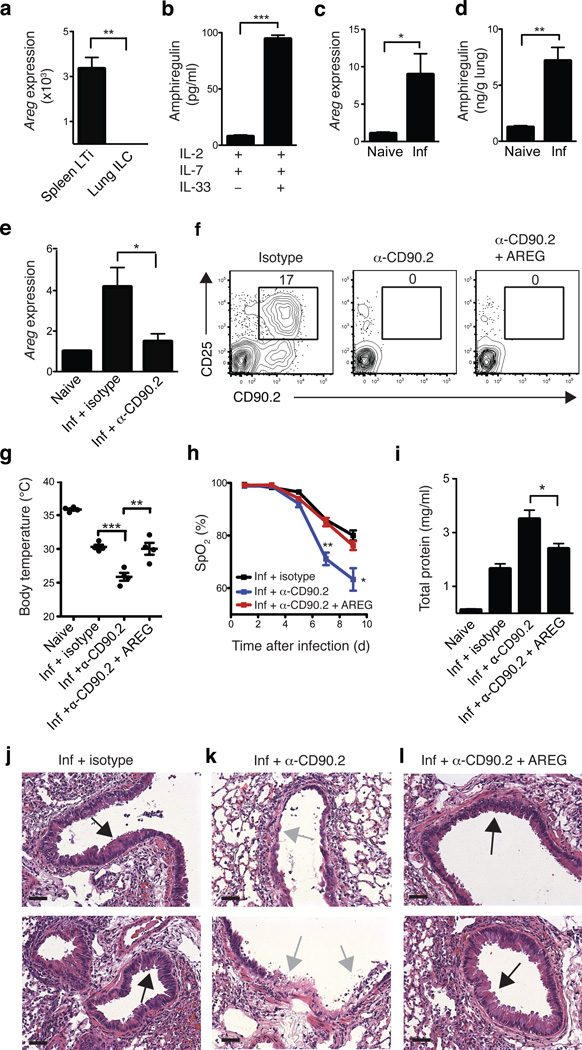
Innate lymphoid cells (ILCs), a heterogeneous cell population, are critical in orchestrating immunity and inflammation in the intestine, but whether ILCs influence immune responses or tissue homeostasis at other mucosal sites remains poorly characterized. Here we identify a population of lung-resident ILCs in mice and humans that expressed the alloantigen Thy-1 (CD90), interleukin 2 (IL-2) receptor a-chain (CD25), IL-7 receptor a-chain (CD127) and the IL-33 receptor subunit T1-ST2. Notably, mouse ILCs accumulated in the lung after infection with influenza virus, and depletion of ILCs resulted in loss of airway epithelial integrity, diminished lung function and impaired airway remodeling. These defects were restored by administration of the lung ILC product amphiregulin. Collectively, our results demonstrate a critical role for lung ILCs in restoring airway epithelial integrity and tissue homeostasis after infection with influenza virus.
Figures
Comment in
-
Innate lymphoid cells wield a double-edged sword.Nat Immunol. 2011 Oct 19;12(11):1025-7. doi: 10.1038/ni.2142. Nat Immunol. 2011. PMID: 22012433 No abstract available.
-
Pulmonary innate lymphoid cells regulate repair and remodeling after acute lung injury.Expert Rev Anti Infect Ther. 2012 Mar;10(3):281-4. doi: 10.1586/eri.12.15. Expert Rev Anti Infect Ther. 2012. PMID: 22397561
References
-
- Turner JR. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol. 2009;9:799–809. - PubMed
-
- Artis D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol. 2008;8:411–420. - PubMed
-
- Sonnenberg GF, Fouser LA, Artis D. Functional biology of the IL-22-IL-22R pathway in regulating immunity and inflammation at barrier surfaces. Adv Immunol. 2010;107:1–29. - PubMed
-
- Spits H, Di Santo JP. The expanding family of innate lymphoid cells: regulators and effectors of immunity and tissue remodeling. Nat Immunol. 2011;12:21–27. - PubMed
-
- Saenz SA, Noti M, Artis D. Innate immune cell populations function as initiators and effectors in Th2 cytokine responses. Trends Immunol. 2010;31:407–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U19 AI083022/AI/NIAID NIH HHS/United States
- R01 AI095466/AI/NIAID NIH HHS/United States
- AI061570/AI/NIAID NIH HHS/United States
- AI091759/AI/NIAID NIH HHS/United States
- AI087990/AI/NIAID NIH HHS/United States
- U01 HL098957/HL/NHLBI NIH HHS/United States
- R37 AI067545/AI/NIAID NIH HHS/United States
- K08 DK093784/DK/NIDDK NIH HHS/United States
- U01 AI095608/AI/NIAID NIH HHS/United States
- R01 AI061570/AI/NIAID NIH HHS/United States
- AI095466/AI/NIAID NIH HHS/United States
- R01 AI074878/AI/NIAID NIH HHS/United States
- U19AI083022/AI/NIAID NIH HHS/United States
- R01 AI091759/AI/NIAID NIH HHS/United States
- DK50306/DK/NIDDK NIH HHS/United States
- T32 AI007532/AI/NIAID NIH HHS/United States
- HL098957/HL/NHLBI NIH HHS/United States
- R01 AI071309/AI/NIAID NIH HHS/United States
- P30 AI045008/AI/NIAID NIH HHS/United States
- AI074878/AI/NIAID NIH HHS/United States
- T32AI007532/AI/NIAID NIH HHS/United States
- R21 AI087990/AI/NIAID NIH HHS/United States
- R01 AI072117/AI/NIAID NIH HHS/United States
- AI095608/AI/NIAID NIH HHS/United States
- T32AI05528/AI/NIAID NIH HHS/United States
- U01AI095608/AI/NIAID NIH HHS/United States
- AI071309/AI/NIAID NIH HHS/United States
- HHSN266200500030C/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
