

Prostaglandin E2 and T cells: friends or foes?
- PMID: 21946663
- PMCID: PMC3389798
- DOI: 10.1038/icb.2011.75
Prostaglandin E2 and T cells: friends or foes?
Abstract
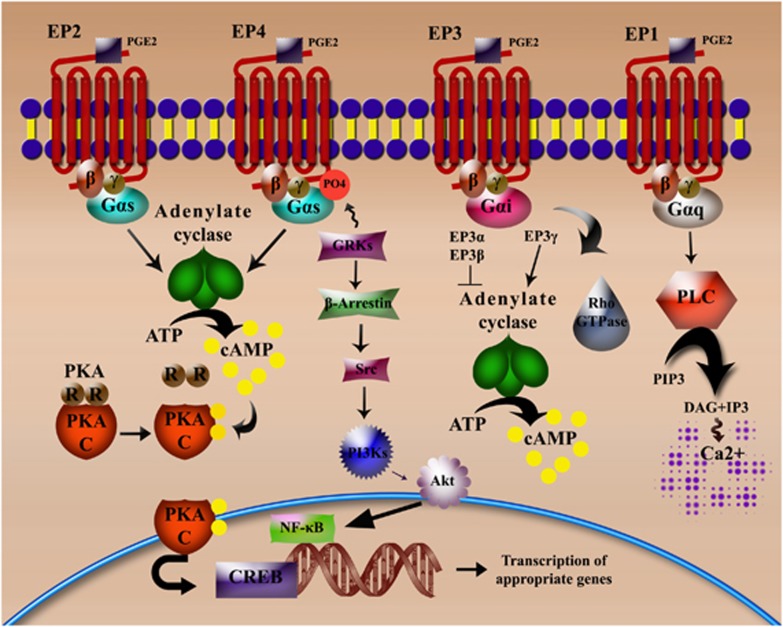
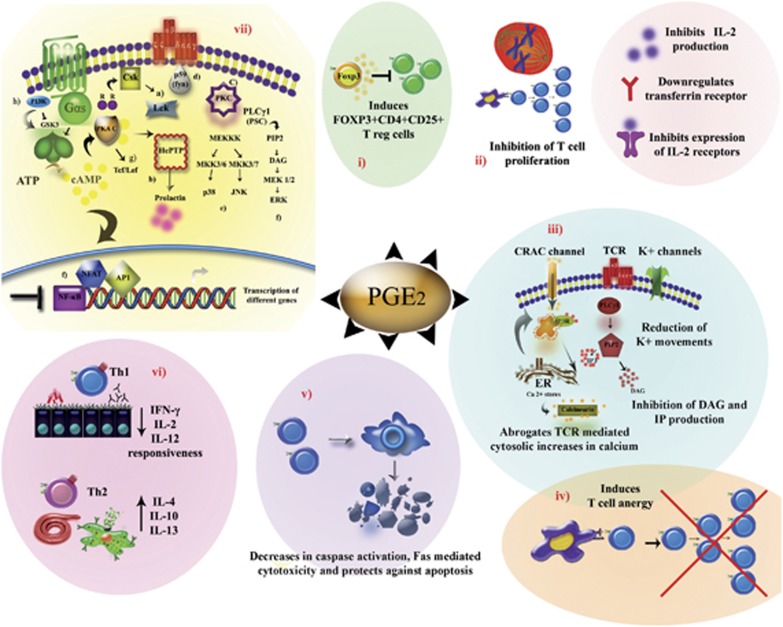
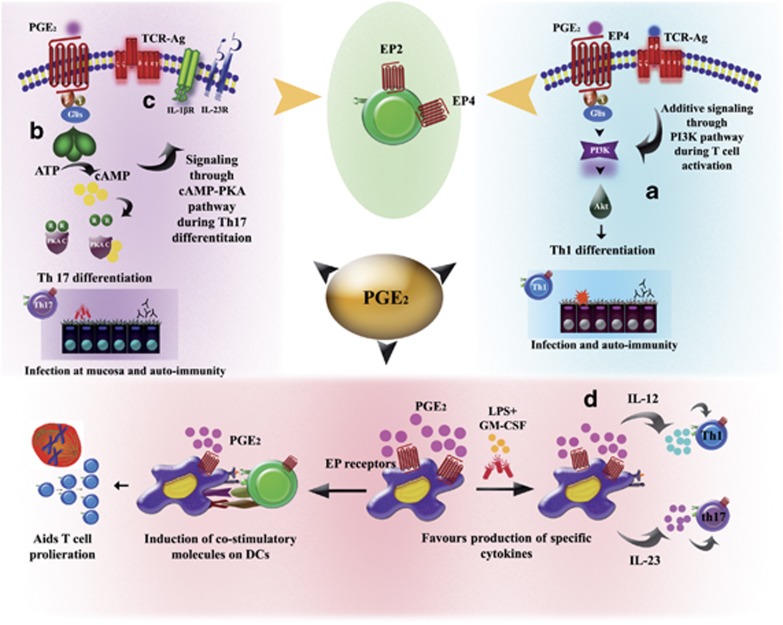
Our understanding of the key players involved in the differential regulation of T-cell responses during inflammation, infection and auto-immunity is fundamental for designing efficient therapeutic strategies against immune diseases. With respect to this, the inhibitory role of the lipid mediator prostaglandin E(2) (PGE(2)) in T-cell immunity has been documented since the 1970s. Studies that ensued investigating the underlying mechanisms substantiated the suppressive function of micromolar concentrations of PGE(2) in T-cell activation, proliferation, differentiation and migration. However, the past decade has seen a revolution in this perspective, since nanomolar concentrations of PGE(2) have been shown to potentiate Th1 and Th17 responses and aid in T-cell proliferation. The understanding of concentration-specific effects of PGE(2) in other cell types, the development of mice deficient in each subtype of the PGE(2) receptors (EP receptors) and the delineation of signalling pathways mediated by the EP receptors have enhanced our understanding of PGE(2) as an immune-stimulator. PGE(2) regulates a multitude of functions in T-cell activation and differentiation and these effects vary depending on the micro-environment of the cell, maturation and activation state of the cell, type of EP receptor involved, local concentration of PGE(2) and whether it is a homeostatic or inflammatory scenario. In this review, we compartmentalize the various aspects of this complex relationship of PGE(2) with T lymphocytes. Given the importance of this molecule in T-cell activation, we also address the possibility of using EP receptor antagonism as a potential therapeutic approach for some immune disorders.
Figures
Similar articles
-
Receptors for prostaglandin E(2) that regulate cellular immune responses in the mouse.J Clin Invest. 2001 Oct;108(8):1229-35. doi: 10.1172/JCI13640. J Clin Invest. 2001. PMID: 11602631 Free PMC article.
-
Stimulation of renin release by prostaglandin E2 is mediated by EP2 and EP4 receptors in mouse kidneys.Am J Physiol Renal Physiol. 2004 Sep;287(3):F427-33. doi: 10.1152/ajprenal.00072.2004. Epub 2004 Apr 27. Am J Physiol Renal Physiol. 2004. PMID: 15113745
-
Prostaglandin E2 modulates the functional responsiveness of human monocytes to chemokines.Eur J Immunol. 2004 Dec;34(12):3682-9. doi: 10.1002/eji.200425226. Eur J Immunol. 2004. PMID: 15484190
-
Prostaglandin E2-induced inflammation: Relevance of prostaglandin E receptors.Biochim Biophys Acta. 2015 Apr;1851(4):414-21. doi: 10.1016/j.bbalip.2014.07.008. Epub 2014 Jul 17. Biochim Biophys Acta. 2015. PMID: 25038274 Review.
-
Roles of the prostaglandin E2 receptors EP subtypes in Alzheimer's disease.Neurosci Bull. 2010 Feb;26(1):77-84. doi: 10.1007/s12264-010-0703-z. Neurosci Bull. 2010. PMID: 20101275 Free PMC article. Review.
Cited by
-
Immune checkpoint blockade in pancreatic cancer: Trudging through the immune desert.Semin Cancer Biol. 2022 Nov;86(Pt 2):14-27. doi: 10.1016/j.semcancer.2022.08.009. Epub 2022 Aug 27. Semin Cancer Biol. 2022. PMID: 36041672 Free PMC article. Review.
-
Targeting cancer with kinase inhibitors.J Clin Invest. 2015 May;125(5):1780-9. doi: 10.1172/JCI76094. Epub 2015 May 1. J Clin Invest. 2015. PMID: 25932675 Free PMC article. Review.
-
Regulatory role of vitamin E in the immune system and inflammation.IUBMB Life. 2019 Apr;71(4):487-494. doi: 10.1002/iub.1976. Epub 2018 Nov 30. IUBMB Life. 2019. PMID: 30501009 Free PMC article.
-
Decreased RORC-dependent silencing of prostaglandin receptor EP2 induces autoimmune Th17 cells.J Clin Invest. 2014 Jun;124(6):2513-22. doi: 10.1172/JCI72973. Epub 2014 May 8. J Clin Invest. 2014. PMID: 24812667 Free PMC article.
-
Tumor-Induced T Cell Polarization by Schwann Cells.Cells. 2022 Nov 9;11(22):3541. doi: 10.3390/cells11223541. Cells. 2022. PMID: 36428970 Free PMC article.
References
-
- Smith WL, Dewitt DL. Prostaglandin endoperoxide H synthases-1 and -2. Adv Immunol. 1996;62:167–215. - PubMed
-
- Harris SG, Padilla J, Koumas L, Ray D, Phipps RP. Prostaglandins as modulators of immunity. Trends Immunol. 2002;23:144–150. - PubMed
-
- Samuelsson B, Goldyne M, Granstrom E, Hamberg M, Hammarstrom S, Malmsten C. Prostaglandins and thromboxanes. Annu Rev Biochem. 1978;47:997–1029. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
