

Investigations on the RNA binding and phosphorylation of groundnut bud necrosis virus nucleocapsid protein
- PMID: 21947504
- PMCID: PMC7086702
- DOI: 10.1007/s00705-011-1110-0
Investigations on the RNA binding and phosphorylation of groundnut bud necrosis virus nucleocapsid protein
Abstract
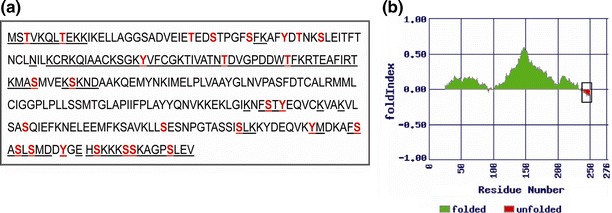
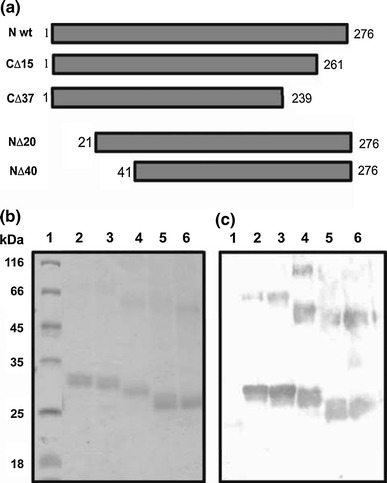
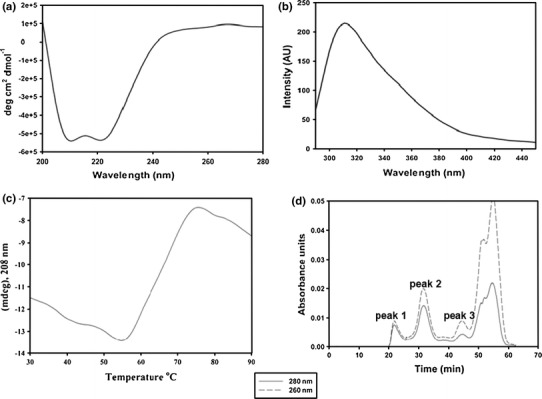
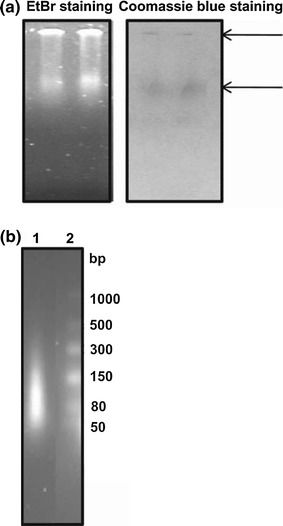
Groundnut bud necrosis virus belongs to the genus Tospovirus, infects a wide range of crop plants and causes severe losses. To understand the role of the nucleocapsid protein in the viral life cycle, the protein was overexpressed in E. coli and purified by Ni-NTA chromatography. The purified N protein was well folded and was predominantly alpha-helical. Deletion analysis revealed that the C-terminal unfolded region of the N protein was involved in RNA binding. Furthermore, the N protein could be phosphorylated in vitro by Nicotiana benthamiana plant sap and by purified recombinant kinases such as protein kinase CK2 and calcium-dependent protein kinase. This is the first report of phoshphorylation of a nucleocapsid protein in the family Bunyaviridae. The possible implications of the present findings for the viral life cycle are discussed.
Figures

Similar articles
-
Characterization of the nucleic acid binding properties of tomato spotted wilt virus nucleocapsid protein.Virology. 1998 Aug 15;248(1):6-11. doi: 10.1006/viro.1998.9223. Virology. 1998. PMID: 9705250
-
Characterization of the large (L) RNA of peanut bud necrosis tospovirus.Arch Virol. 1998;143(12):2381-90. doi: 10.1007/s007050050468. Arch Virol. 1998. PMID: 9930194 Free PMC article.
-
Asymmetric Trimeric Ring Structure of the Nucleocapsid Protein of Tospovirus.J Virol. 2017 Sep 27;91(20):e01002-17. doi: 10.1128/JVI.01002-17. Print 2017 Oct 15. J Virol. 2017. PMID: 28768868 Free PMC article.
-
The nucleocapsid of vesicular stomatitis virus.Sci China Life Sci. 2012 Apr;55(4):291-300. doi: 10.1007/s11427-012-4307-x. Epub 2012 May 9. Sci China Life Sci. 2012. PMID: 22566085 Review.
-
Nucleic acid chaperone activity of HIV-1 nucleocapsid protein: critical role in reverse transcription and molecular mechanism.Prog Nucleic Acid Res Mol Biol. 2005;80:217-86. doi: 10.1016/S0079-6603(05)80006-6. Prog Nucleic Acid Res Mol Biol. 2005. PMID: 16164976 Review. No abstract available.
Cited by
-
Two key arginine residues in the coat protein of Bamboo mosaic virus differentially affect the accumulation of viral genomic and subgenomic RNAs.Mol Plant Pathol. 2014 Feb;15(2):196-210. doi: 10.1111/mpp.12080. Epub 2013 Oct 28. Mol Plant Pathol. 2014. PMID: 24393453 Free PMC article.
-
Five questions on the cell-to-cell movement of Orthotospoviruses.BBA Adv. 2024 Oct 16;6:100124. doi: 10.1016/j.bbadva.2024.100124. eCollection 2024. BBA Adv. 2024. PMID: 39498475 Free PMC article.
-
Phosphorylation regulates the subcellular localization of Cucumber Mosaic Virus 2b protein.Sci Rep. 2017 Oct 18;7(1):13444. doi: 10.1038/s41598-017-13870-7. Sci Rep. 2017. PMID: 29044170 Free PMC article.
-
Raman-dielectrophoresis goes viral: towards a rapid and label-free platform for plant virus characterization.Front Microbiol. 2023 Nov 22;14:1292461. doi: 10.3389/fmicb.2023.1292461. eCollection 2023. Front Microbiol. 2023. PMID: 38075896 Free PMC article.
-
Phosphorylation of Beet black scorch virus coat protein by PKA is required for assembly and stability of virus particles.Sci Rep. 2015 Jun 25;5:11585. doi: 10.1038/srep11585. Sci Rep. 2015. PMID: 26108567 Free PMC article.
References
-
- Chang CK, Hsu YL, Chang YH, Chao FA, Wu MC, Huang YS, Hu CK, Huang TH. Multiple nucleic acid binding sites and intrinsic disorder of severe acute respiratory syndrome coronavirus nucleocapsid protein: implications for ribonucleocapsid protein packaging. J Virol. 2009;83:2255–2264. doi: 10.1128/JVI.02001-08. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
