

Post-transcriptional exon shuffling events in humans can be evolutionarily conserved and abundant
- PMID: 21948523
- PMCID: PMC3205564
- DOI: 10.1101/gr.116442.110
Post-transcriptional exon shuffling events in humans can be evolutionarily conserved and abundant
Abstract
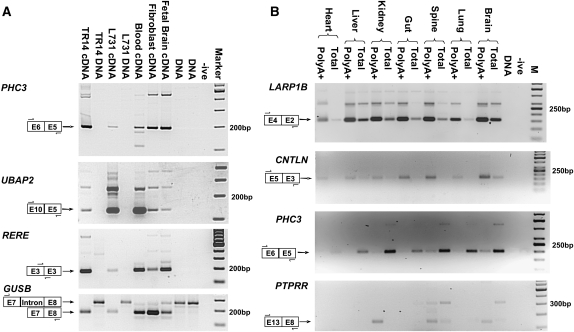
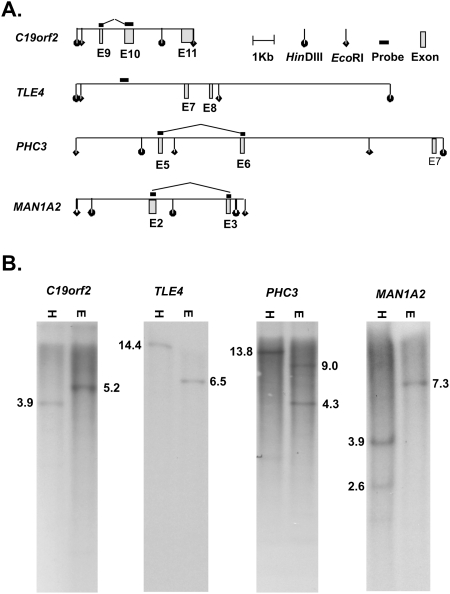
In silico analyses have established that transcripts from some genes can be processed into RNAs with rearranged exon order relative to genomic structure (post-transcriptional exon shuffling, or PTES). Although known to contribute to transcriptome diversity in some species, to date the structure, distribution, abundance, and functional significance of human PTES transcripts remains largely unknown. Here, using high-throughput transcriptome sequencing, we identify 205 putative human PTES products from 176 genes. We validate 72 out of 112 products analyzed using RT-PCR, and identify additional PTES products structurally related to 61% of validated targets. Sequencing of these additional products reveals GT-AG dinucleotides at >95% of the splice junctions, confirming that they are processed by the spliceosome. We show that most PTES transcripts are expressed in a wide variety of human tissues, that they can be polyadenylated, and that some are conserved in mouse. We also show that they can extend into 5' and 3' UTRs, consistent with formation via trans-splicing of independent pre-mRNA molecules. Finally, we use real-time PCR to compare the abundance of PTES exon junctions relative to canonical exon junctions within the transcripts from seven genes. PTES exon junctions are present at <0.01% to >90% of the levels of canonical junctions, with transcripts from MAN1A2, PHC3, TLE4, and CDK13 exhibiting the highest levels. This is the first systematic experimental analysis of PTES in human, and it suggests both that the phenomenon is much more widespread than previously thought and that some PTES transcripts could be functional.
Figures




References
-
- Akopian AN, Okuse K, Souslova V, England S, Ogata N, Wood JN 1999. Trans-splicing of a voltage-gated sodium channel is regulated by nerve growth factor. FEBS Lett 445: 177–182 - PubMed
-
- Blumenthal T 1995. Trans-splicing and polycistronic transcription in Ceanorhabditis elegans. Trends Genet 11: 132–136 - PubMed
-
- Case M, Matheson E, Minto L, Hassan R, Harrison CJ, Bown N, Bailey S, Vormoor J, Hall AG, Irving JAE 2008. Mutation of genes impacting on the RAS pathway are common in childhood acute lymphoblastic leukemia. Cancer Res 68: 6803–6809 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous