

Whole-genome association study on tissue tropism phenotypes in group A Streptococcus
- PMID: 21949075
- PMCID: PMC3232895
- DOI: 10.1128/JB.05263-11
Whole-genome association study on tissue tropism phenotypes in group A Streptococcus
Abstract
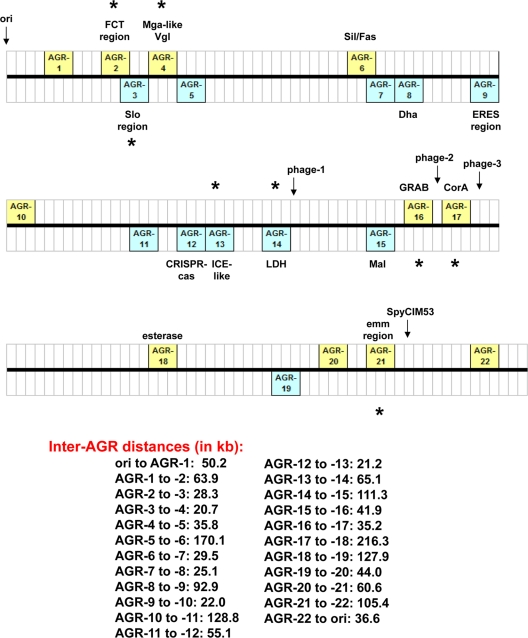
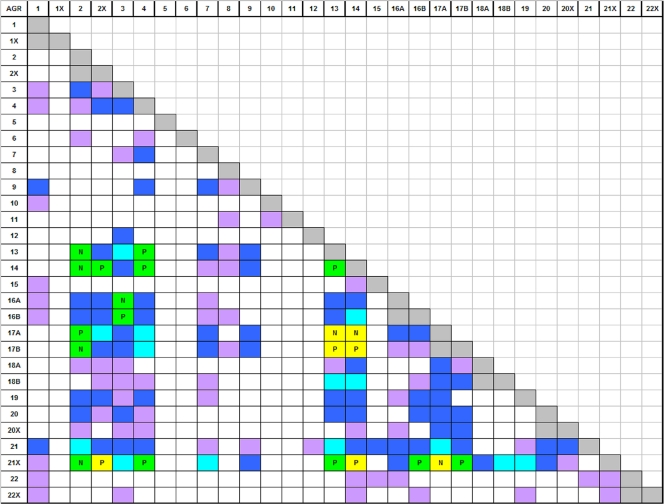
Group A Streptococcus (GAS) has a rich evolutionary history of horizontal transfer among its core genes. Yet, despite extensive genetic mixing, GAS strains have discrete ecological phenotypes. To further our understanding of the molecular basis for ecological phenotypes, comparative genomic hybridization of a set of 97 diverse strains to a GAS pangenome microarray was undertaken, and the association of accessory genes with emm genotypes that define tissue tropisms for infection was determined. Of the 22 nonprophage accessory gene regions (AGRs) identified, only 3 account for all statistically significant linkage disequilibrium among strains having the genotypic biomarkers for throat versus skin infection specialists. Networked evolution and population structure analyses of loci representing each of the AGRs reveal that most strains with the skin specialist and generalist biomarkers form discrete clusters, whereas strains with the throat specialist biomarker are highly diverse. To identify coinherited and coselected accessory genes, the strength of genetic associations was determined for all possible pairwise combinations of accessory genes among the 97 GAS strains. Accessory genes showing very strong associations provide the basis for an evolutionary model, which reveals that a major transition between many throat and skin specialist haplotypes correlates with the gain or loss of genes encoding fibronectin-binding proteins. This study employs a novel synthesis of tools to help delineate the major genetic changes associated with key adaptive shifts in an extensively recombined bacterial species.
Figures





Similar articles
-
Comparative analysis of emm type pattern of Group A Streptococcus throat and skin isolates from India and their association with closely related SIC, a streptococcal virulence factor.BMC Microbiol. 2008 Sep 16;8:150. doi: 10.1186/1471-2180-8-150. BMC Microbiol. 2008. PMID: 18796133 Free PMC article.
-
Natural selection and evolution of streptococcal virulence genes involved in tissue-specific adaptations.J Bacteriol. 2004 Jan;186(1):110-21. doi: 10.1128/JB.186.1.110-121.2004. J Bacteriol. 2004. PMID: 14679231 Free PMC article.
-
Evolution of transcription regulatory genes is linked to niche specialization in the bacterial pathogen Streptococcus pyogenes.J Bacteriol. 2005 Jun;187(12):4163-72. doi: 10.1128/JB.187.12.4163-4172.2005. J Bacteriol. 2005. PMID: 15937178 Free PMC article.
-
Tissue tropisms in group A streptococcal infections.Future Microbiol. 2010 Apr;5(4):623-38. doi: 10.2217/fmb.10.28. Future Microbiol. 2010. PMID: 20353302 Free PMC article. Review.
-
Tissue tropisms in group A Streptococcus: what virulence factors distinguish pharyngitis from impetigo strains?Curr Opin Infect Dis. 2016 Jun;29(3):295-303. doi: 10.1097/QCO.0000000000000262. Curr Opin Infect Dis. 2016. PMID: 26895573 Free PMC article. Review.
Cited by
-
Population genomics: an investigative tool for epidemics.Am J Pathol. 2012 Apr;180(4):1358-61. doi: 10.1016/j.ajpath.2012.01.007. Epub 2012 Feb 28. Am J Pathol. 2012. PMID: 22386771 Free PMC article.
-
Concerns for efficacy of a 30-valent M-protein-based Streptococcus pyogenes vaccine in regions with high rates of rheumatic heart disease.PLoS Negl Trop Dis. 2019 Jul 3;13(7):e0007511. doi: 10.1371/journal.pntd.0007511. eCollection 2019 Jul. PLoS Negl Trop Dis. 2019. PMID: 31269021 Free PMC article.
-
Incremental Contributions of FbaA and Other Impetigo-Associated Surface Proteins to Fitness and Virulence of a Classical Group A Streptococcal Skin Strain.Infect Immun. 2017 Oct 18;85(11):e00374-17. doi: 10.1128/IAI.00374-17. Print 2017 Nov. Infect Immun. 2017. PMID: 28808160 Free PMC article.
-
Parasite spillover rather than niche expansion explains infection of host brain by diplostomid eye flukes.Proc Biol Sci. 2025 Feb;292(2040):20242648. doi: 10.1098/rspb.2024.2648. Epub 2025 Feb 5. Proc Biol Sci. 2025. PMID: 39904393 Free PMC article.
-
Invasive group a streptococcal infection associated with community healthcare services delivered at home, South East England, December 2021-2023: Descriptive epidemiological study.Epidemiol Infect. 2025 Mar 21;153:e56. doi: 10.1017/S0950268825000287. Epidemiol Infect. 2025. PMID: 40114514 Free PMC article.
References
-
- Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403–410 - PubMed
-
- Anthony B. F., Kaplan E. L., Wannamaker L. W., Chapman S. S. 1976. The dynamics of streptococcal infections in a defined population of children: serotypes associated with skin and respiratory infections. Am. J. Epidemiol. 104:652–666 - PubMed
-
- Banks D. J., Porcella S. F., Barbian K. D., Martin J. M., Musser J. M. 2003. Structure and distribution of an unusual chimeric genetic element encoding macrolide resistance in phylogenetically diverse clones of group A Streptococcus. J. Infect. Dis. 188:1898–1908 - PubMed
-
- Beall B. 16 May 2011, posting date Streptococcus pyogenes emm sequence database. http://cdc.gov/ncidod/biotech/strep/strepblast.htm
-
- Bendtsen J. D., Nielsen H., von Heijne G., Brunak S. 2004. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340:783–795 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
