

VasH is a transcriptional regulator of the type VI secretion system functional in endemic and pandemic Vibrio cholerae
- PMID: 21949076
- PMCID: PMC3232897
- DOI: 10.1128/JB.05414-11
VasH is a transcriptional regulator of the type VI secretion system functional in endemic and pandemic Vibrio cholerae
Abstract
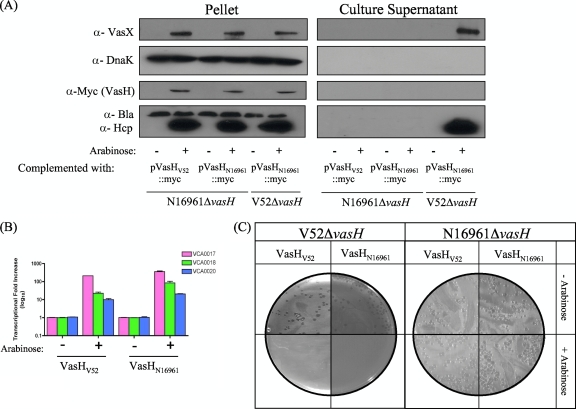
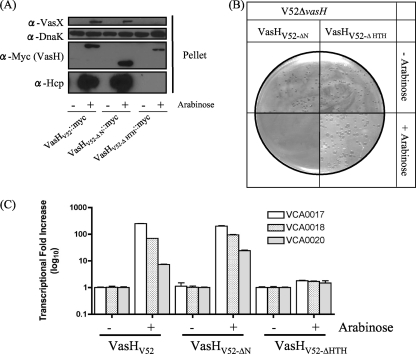
The gram-negative bacterium Vibrio cholerae is the etiological agent of cholera, a disease characterized by the release of high volumes of watery diarrhea. Many medically important proteobacteria, including V. cholerae, carry one or multiple copies of the gene cluster that encodes the bacterial type VI secretion system (T6SS) to confer virulence or interspecies competitiveness. Structural similarity and sequence homology between components of the T6SS and the cell-puncturing device of T4 bacteriophage suggest that the T6SS functions as a molecular syringe to inject effector molecules into prokaryotic and eukaryotic target cells. Although our understanding of how the structural T6SS apparatus assembles is developing, little is known about how this system is regulated. Here, we report on the contribution of the activator of the alternative sigma factor 54, VasH, as a global regulator of the V. cholerae T6SS. Using bioinformatics and mutational analyses, we identified domains of the VasH polypeptide that are essential for its ability to initiate transcription of T6SS genes and established a universal role for VasH in endemic and pandemic V. cholerae strains.
Figures





Similar articles
-
Constitutive type VI secretion system expression gives Vibrio cholerae intra- and interspecific competitive advantages.PLoS One. 2012;7(10):e48320. doi: 10.1371/journal.pone.0048320. Epub 2012 Oct 26. PLoS One. 2012. PMID: 23110230 Free PMC article.
-
Vibrio cholerae requires the type VI secretion system virulence factor VasX to kill Dictyostelium discoideum.Infect Immun. 2011 Jul;79(7):2941-9. doi: 10.1128/IAI.01266-10. Epub 2011 May 9. Infect Immun. 2011. PMID: 21555399 Free PMC article.
-
The Vibrio cholerae type VI secretion system employs diverse effector modules for intraspecific competition.Nat Commun. 2014 Apr 1;5:3549. doi: 10.1038/ncomms4549. Nat Commun. 2014. PMID: 24686479 Free PMC article.
-
The Vibrio cholerae type VI secretion system: toxins, regulators and consequences.Environ Microbiol. 2020 Oct;22(10):4112-4122. doi: 10.1111/1462-2920.14976. Epub 2020 Mar 13. Environ Microbiol. 2020. PMID: 32133757 Review.
-
Rules of Engagement: The Type VI Secretion System in Vibrio cholerae.Trends Microbiol. 2017 Apr;25(4):267-279. doi: 10.1016/j.tim.2016.12.003. Epub 2016 Dec 24. Trends Microbiol. 2017. PMID: 28027803 Free PMC article. Review.
Cited by
-
Structure and regulation of the type VI secretion system.Annu Rev Microbiol. 2012;66:453-72. doi: 10.1146/annurev-micro-121809-151619. Epub 2012 Jun 28. Annu Rev Microbiol. 2012. PMID: 22746332 Free PMC article. Review.
-
A view to a kill: the bacterial type VI secretion system.Cell Host Microbe. 2014 Jan 15;15(1):9-21. doi: 10.1016/j.chom.2013.11.008. Epub 2013 Dec 11. Cell Host Microbe. 2014. PMID: 24332978 Free PMC article. Review.
-
The LonA Protease Regulates Biofilm Formation, Motility, Virulence, and the Type VI Secretion System in Vibrio cholerae.J Bacteriol. 2016 Jan 11;198(6):973-85. doi: 10.1128/JB.00741-15. J Bacteriol. 2016. PMID: 26755629 Free PMC article.
-
Sensing of intracellular Hcp levels controls T6SS expression in Vibrio cholerae.Proc Natl Acad Sci U S A. 2021 Jun 22;118(25):e2104813118. doi: 10.1073/pnas.2104813118. Proc Natl Acad Sci U S A. 2021. PMID: 34161288 Free PMC article.
-
Proteus mirabilis Employs a Contact-Dependent Killing System against Competing Enterobacteriaceae.mSphere. 2021 Aug 25;6(4):e0032121. doi: 10.1128/mSphere.00321-21. Epub 2021 Jul 28. mSphere. 2021. PMID: 34319125 Free PMC article.
References
-
- Barrett J., Ray P., Sobczyk A., Little R., Dixon R. 2001. Concerted inhibition of the transcriptional activation functions of the enhancer-binding protein NIFA by the anti-activator NIFL. Mol. Microbiol. 39:480–493 - PubMed
-
- Bhattacharya M. K., et al. 1993. Outbreak of Vibrio cholerae non-O1 in India and Bangladesh. Lancet 341:1346–1347 - PubMed
-
- Colwell R. R. 1996. Global climate and infectious disease: the cholera paradigm. Science 274:2025–2031 - PubMed
-
- Cvjetanovic B., Barua D. 1972. The seventh pandemic of cholera. Nature 239:137–138 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
