

Evolutionary remodeling of βγ-crystallins for domain stability at cost of Ca2+ binding
- PMID: 21949186
- PMCID: PMC3243557
- DOI: 10.1074/jbc.M111.247890
Evolutionary remodeling of βγ-crystallins for domain stability at cost of Ca2+ binding
Abstract
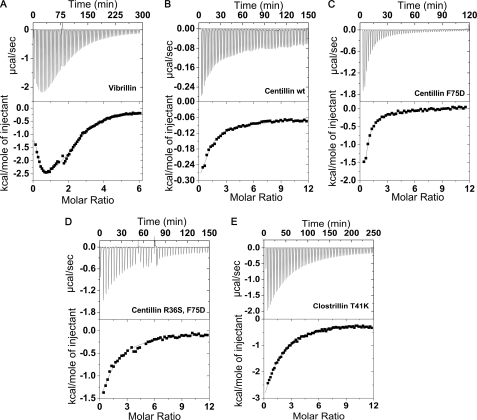
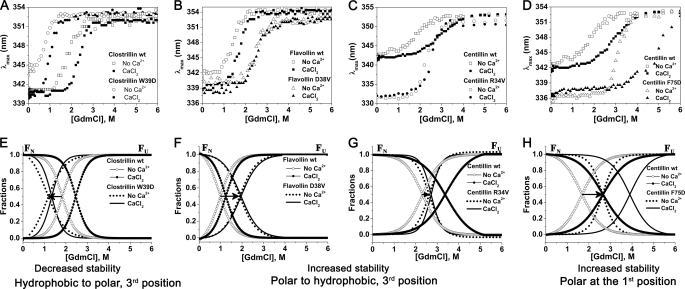
The topologically similar βγ-crystallins that are prevalent in all kingdoms of life have evolved for high innate domain stability to perform their specialized functions. The evolution of stability and its control in βγ-crystallins that possess either a canonical (mostly from microorganisms) or degenerate (principally found in vertebrate homologues) Ca2+-binding motif is not known. Using equilibrium unfolding of βγ-crystallin domains (26 wild-type domains and their mutants) in apo- and holo-forms, we demonstrate the presence of a stability gradient across these members, which is attained by the choice of residues in the (N/D)(N/D)XX(S/T)S Ca2+-binding motif. The occurrence of a polar, hydrophobic, or Ser residue at the 1st, 3rd, or 5th position of the motif is likely linked to a higher domain stability. Partial conversion of a microbe-type domain (with a canonical Ca2+-binding motif) to a vertebrate-type domain (with a degenerate Ca2+-binding motif) by mutating serine to arginine/lysine disables the Ca2+-binding but significantly augments its stability. Conversely, stability is compromised when arginine (in a vertebrate-type disabled domain) is replaced by serine (as a microbe type). Our results suggest that such conversions were acquired as a strategy for desired stability in vertebrate members at the cost of Ca2+-binding. In a physiological context, we demonstrate that a mutation such as an arginine to serine (R77S) mutation in this motif of γ-crystallin (partial conversion to microbe-type), implicated in cataracts, decreases the domain stability. Thus, this motif acts as a "central tuning knob" for innate as well as Ca2+-induced gain in stability, incorporating a stability gradient across βγ-crystallin members critical for their specialized functions.
Figures







Similar articles
-
Ca2+-binding motif of βγ-crystallins.J Biol Chem. 2014 Apr 18;289(16):10958-10966. doi: 10.1074/jbc.O113.539569. Epub 2014 Feb 24. J Biol Chem. 2014. PMID: 24567326 Free PMC article. Review.
-
Association properties and unfolding of a βγ-crystallin domain of a Vibrio-specific protein.PLoS One. 2013;8(1):e53610. doi: 10.1371/journal.pone.0053610. Epub 2013 Jan 22. PLoS One. 2013. PMID: 23349723 Free PMC article.
-
βγ-Crystallination Endows a Novel Bacterial Glycoside Hydrolase 64 with Ca2+-Dependent Activity Modulation.J Bacteriol. 2019 Nov 5;201(23):e00392-19. doi: 10.1128/JB.00392-19. Print 2019 Dec 1. J Bacteriol. 2019. PMID: 31527113 Free PMC article.
-
Molecular evolution of the betagamma lens crystallin superfamily: evidence for a retained ancestral function in gamma N crystallins?Mol Biol Evol. 2009 May;26(5):1127-42. doi: 10.1093/molbev/msp028. Epub 2009 Feb 20. Mol Biol Evol. 2009. PMID: 19233964
-
Microbial βγ-crystallins.Prog Biophys Mol Biol. 2014 Jul;115(1):42-51. doi: 10.1016/j.pbiomolbio.2014.02.007. Epub 2014 Mar 1. Prog Biophys Mol Biol. 2014. PMID: 24594023 Review.
Cited by
-
Divalent Cations and the Divergence of βγ-Crystallin Function.Biochemistry. 2019 Nov 12;58(45):4505-4518. doi: 10.1021/acs.biochem.9b00507. Epub 2019 Nov 1. Biochemistry. 2019. PMID: 31647219 Free PMC article.
-
A mutation in the start codon of γ-crystallin D leads to nuclear cataracts in the Dahl SS/Jr-Ctr strain.Mamm Genome. 2013 Apr;24(3-4):95-104. doi: 10.1007/s00335-013-9447-1. Epub 2013 Feb 13. Mamm Genome. 2013. PMID: 23404175 Free PMC article.
-
The mutation V42M distorts the compact packing of the human gamma-S-crystallin molecule, resulting in congenital cataract.PLoS One. 2012;7(12):e51401. doi: 10.1371/journal.pone.0051401. Epub 2012 Dec 21. PLoS One. 2012. PMID: 23284690 Free PMC article.
-
Functional Role of γ-Crystallin N in the Auditory Hindbrain.PLoS One. 2016 Aug 12;11(8):e0161140. doi: 10.1371/journal.pone.0161140. eCollection 2016. PLoS One. 2016. PMID: 27517863 Free PMC article.
-
Single-molecule Force Spectroscopy Reveals the Calcium Dependence of the Alternative Conformations in the Native State of a βγ-Crystallin Protein.J Biol Chem. 2016 Aug 26;291(35):18263-75. doi: 10.1074/jbc.M116.729525. Epub 2016 Jul 4. J Biol Chem. 2016. PMID: 27378818 Free PMC article.
References
-
- Blundell T., Lindley P., Miller L., Moss D., Slingsby C., Tickle I., Turnell B., Wistow G. (1981) Nature 289, 771–777 - PubMed
-
- Wistow G., Turnell B., Summers L., Slingsby C., Moss D., Miller L., Lindley P., Blundell T. (1983) J. Mol. Biol. 170, 175–202 - PubMed
-
- Jaenicke R. (1994) Naturwissenschaften 81, 423–429 - PubMed
-
- Jaenicke R., Slingsby C. (2001) Crit. Rev. Biochem. Mol. Biol. 36, 435–499 - PubMed
-
- Bloemendal H., de Jong W., Jaenicke R., Lubsen N. H., Slingsby C., Tardieu A. (2004) Prog. Biophys. Mol. Biol. 86, 407–485 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
