

HSP90 functions in the circadian clock through stabilization of the client F-box protein ZEITLUPE
- PMID: 21949396
- PMCID: PMC3189077
- DOI: 10.1073/pnas.1110406108
HSP90 functions in the circadian clock through stabilization of the client F-box protein ZEITLUPE
Abstract
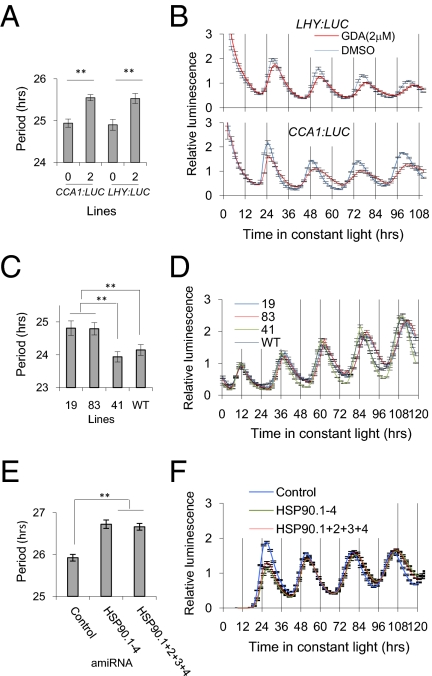
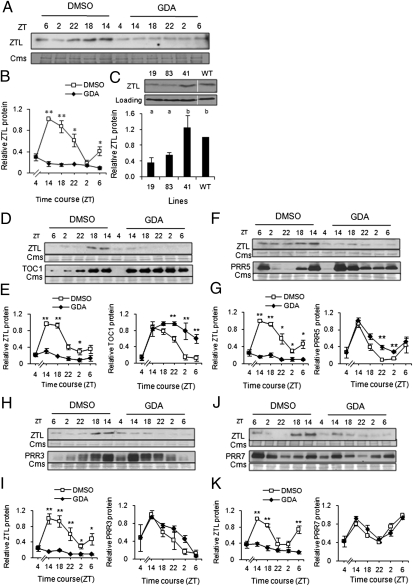
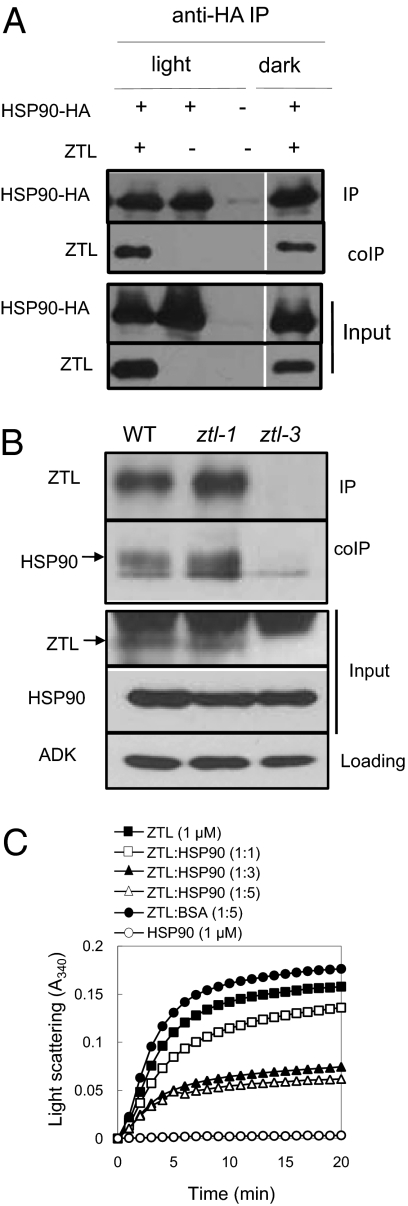
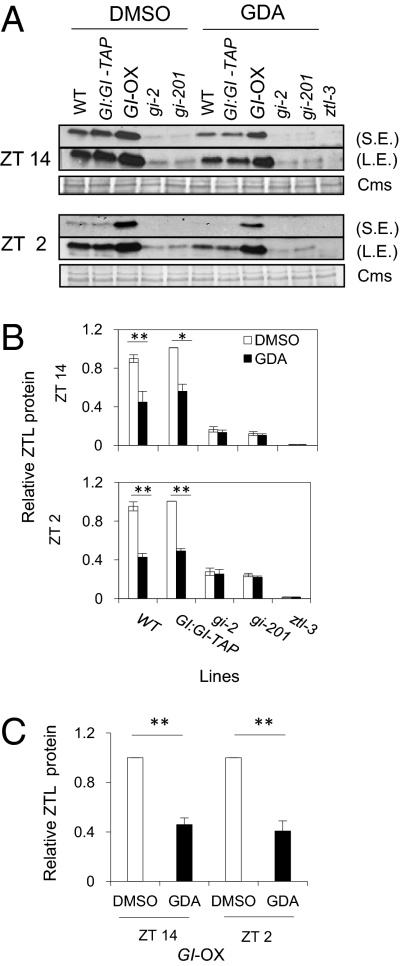
The autoregulatory loops of the circadian clock consist of feedback regulation of transcription/translation circuits but also require finely coordinated cytoplasmic and nuclear proteostasis. Although protein degradation is important to establish steady-state levels, maturation into their active conformation also factors into protein homeostasis. HSP90 facilitates the maturation of a wide range of client proteins, and studies in metazoan clocks implicate HSP90 as an integrator of input or output. Here we show that the Arabidopsis circadian clock-associated F-box protein ZEITLUPE (ZTL) is a unique client for cytoplasmic HSP90. The HSP90-specific inhibitor geldanamycin and RNAi-mediated depletion of cytoplasmic HSP90 reduces levels of ZTL and lengthens circadian period, consistent with ztl loss-of-function alleles. Transient transfection of artificial microRNA targeting cytoplasmic HSP90 genes similarly lengthens period. Proteolytic targets of SCF(ZTL), TOC1 and PRR5, are stabilized in geldanamycin-treated seedlings, whereas the levels of closely related clock proteins, PRR3 and PRR7, are unchanged. An in vitro holdase assay, typically used to demonstrate chaperone activity, shows that ZTL can be effectively bound, and aggregation prevented, by HSP90. GIGANTEA, a unique stabilizer of ZTL, may act in the same pathway as HSP90, possibly linking these two proteins to a similar mechanism. Our findings establish maturation of ZTL by HSP90 as essential for proper function of the Arabidopsis circadian clock. Unlike metazoan systems, HSP90 functions here within the core oscillator. Additionally, F-box proteins as clients may place HSP90 in a unique and more central role in proteostasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Michael TP, et al. Enhanced fitness conferred by naturally occurring variation in the circadian clock. Science. 2003;302:1049–1053. - PubMed
-
- Dodd AN, et al. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science. 2005;309:630–633. - PubMed
-
- Yerushalmi S, Yakir E, Green RM. Circadian clocks and adaptation in Arabidopsis. Mol Ecol. 2011;20:1155–1165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
