

EMILIN1-α4/α9 integrin interaction inhibits dermal fibroblast and keratinocyte proliferation
- PMID: 21949412
- PMCID: PMC3187715
- DOI: 10.1083/jcb.201008013
EMILIN1-α4/α9 integrin interaction inhibits dermal fibroblast and keratinocyte proliferation
Abstract
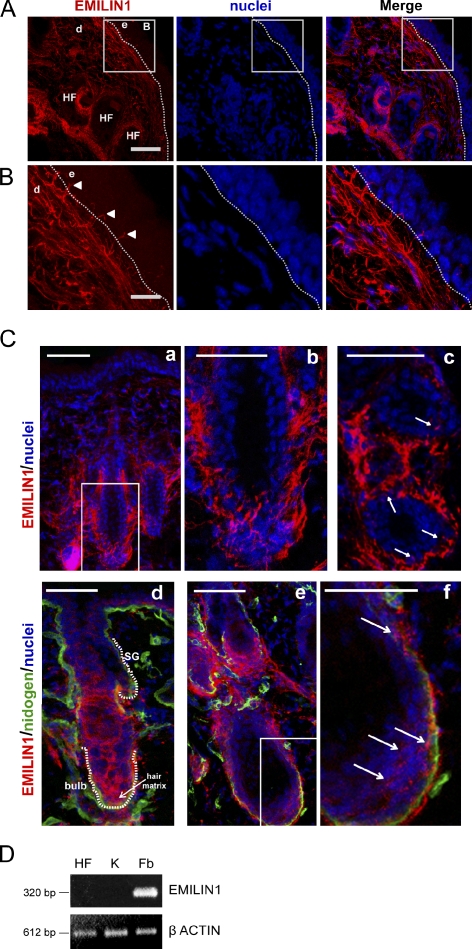
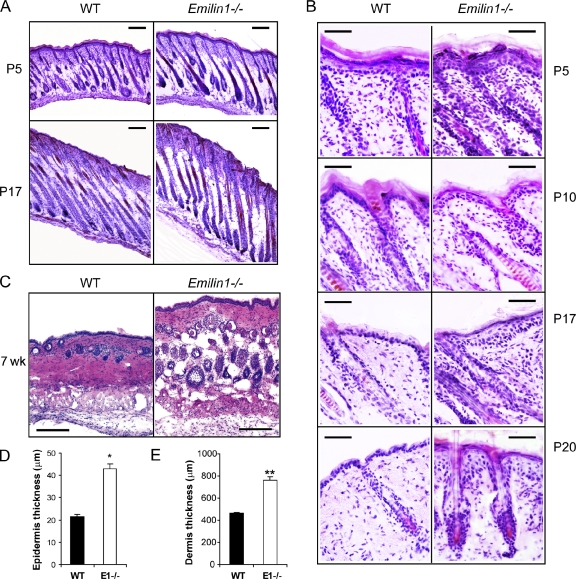
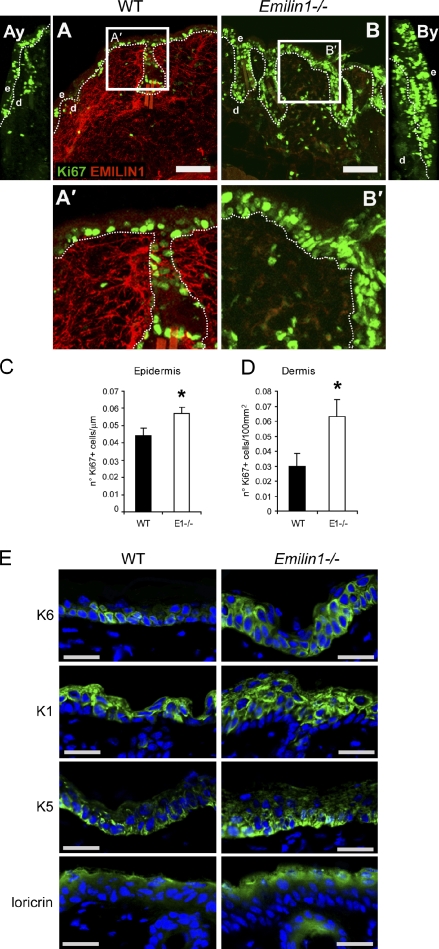
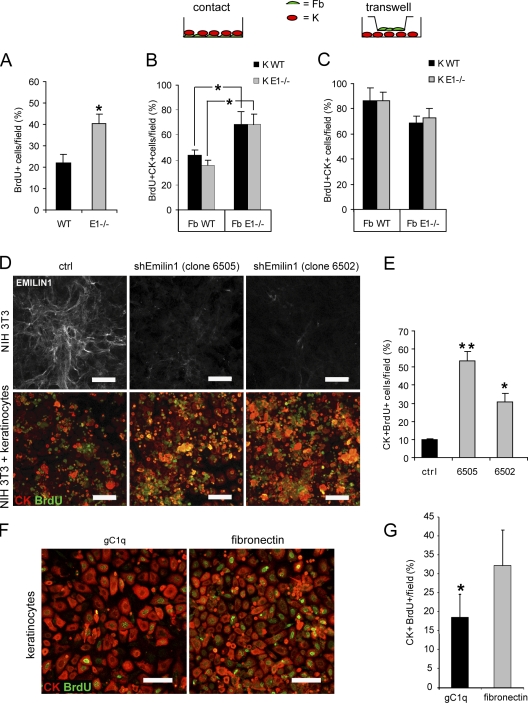
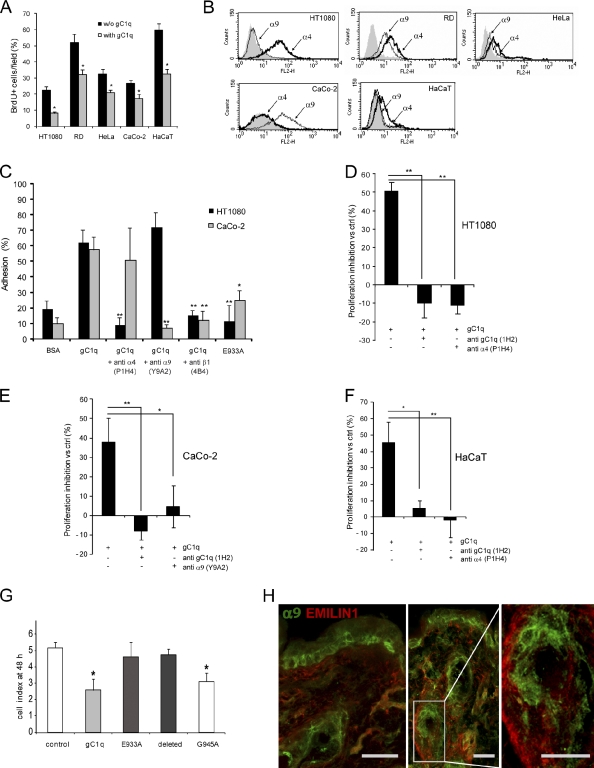
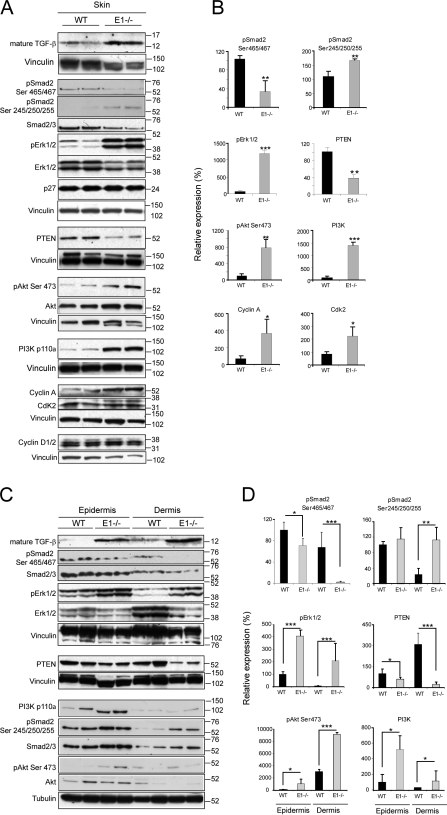
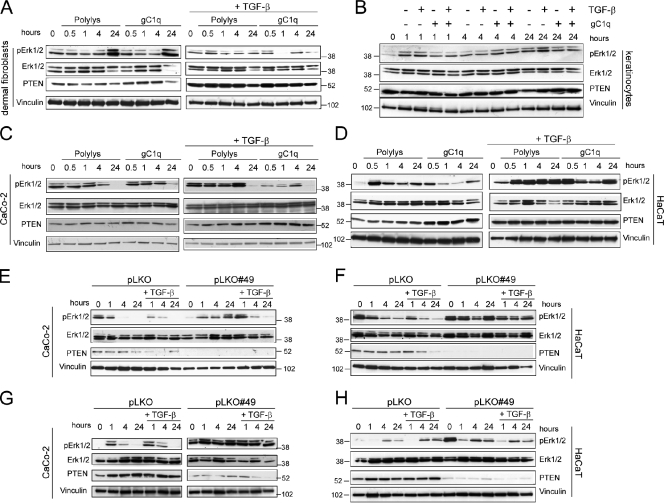
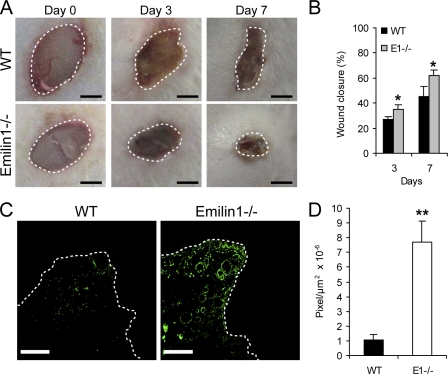
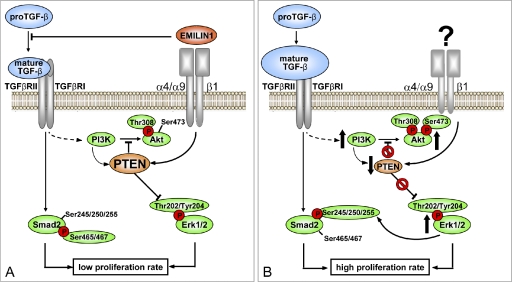
EMILIN1 promotes α4β1 integrin-dependent cell adhesion and migration and reduces pro-transforming growth factor-β processing. A knockout mouse model was used to unravel EMILIN1 functions in skin where the protein was abundantly expressed in the dermal stroma and where EMILIN1-positive fibrils reached the basal keratinocyte layer. Loss of EMILIN1 caused dermal and epidermal hyperproliferation and accelerated wound closure. We identified the direct engagement of EMILIN1 to α4β1 and α9β1 integrins as the mechanism underlying the homeostatic role exerted by EMILIN1. The lack of EMILIN1-α4/α9 integrin interaction was accompanied by activation of PI3K/Akt and Erk1/2 pathways as a result of the reduction of PTEN. The down-regulation of PTEN empowered Erk1/2 phosphorylation that in turn inhibited Smad2 signaling by phosphorylation of residues Ser245/250/255. These results highlight the important regulatory role of an extracellular matrix component in skin proliferation. In addition, EMILIN1 is identified as a novel ligand for keratinocyte α9β1 integrin, suggesting prospective roles for this receptor-ligand pair in skin homeostasis.
Figures
References
-
- Böttinger E.P., Jakubczak J.L., Roberts I.S., Mumy M., Hemmati P., Bagnall K., Merlino G., Wakefield L.M. 1997. Expression of a dominant-negative mutant TGF-beta type II receptor in transgenic mice reveals essential roles for TGF-beta in regulation of growth and differentiation in the exocrine pancreas. EMBO J. 16:2621–2633 10.1093/emboj/16.10.2621 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
