

Regulation of villin by wnt5a/ror2 signaling in human intestinal cells
- PMID: 21949508
- PMCID: PMC3171703
- DOI: 10.3389/fphys.2011.00058
Regulation of villin by wnt5a/ror2 signaling in human intestinal cells
Abstract
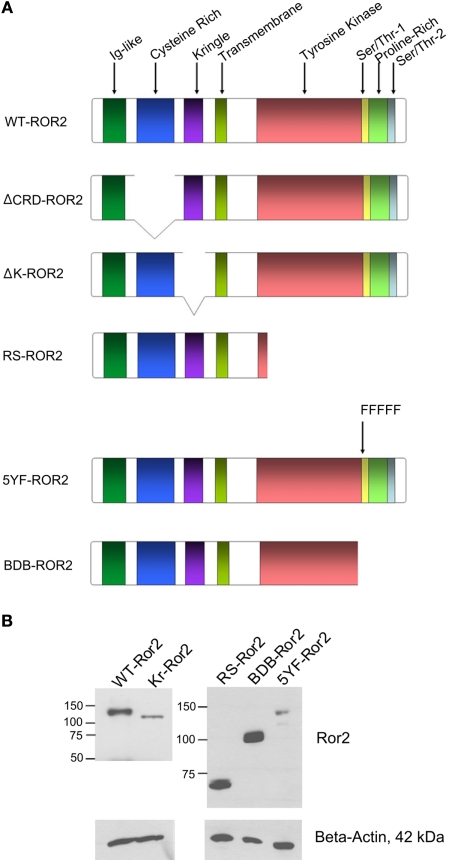
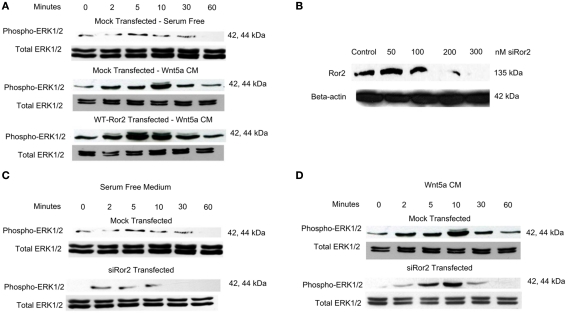
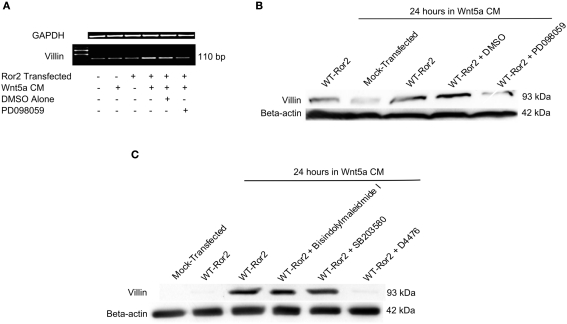
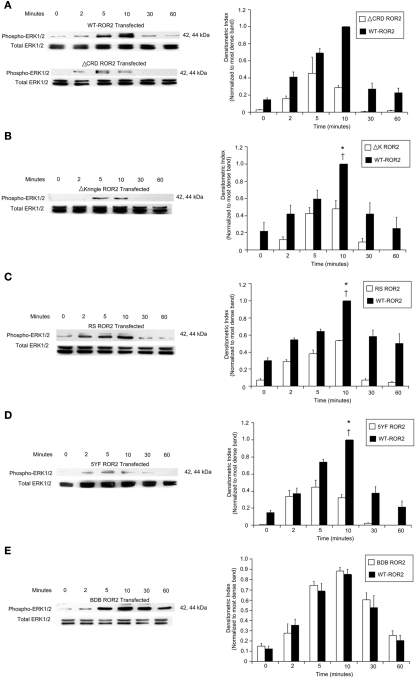
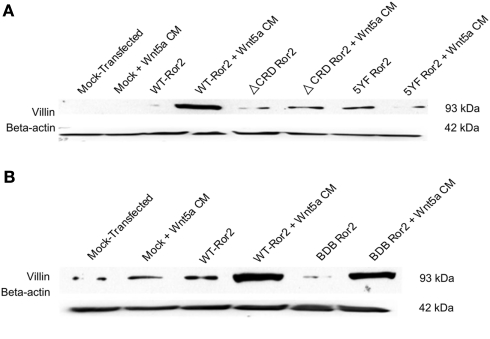
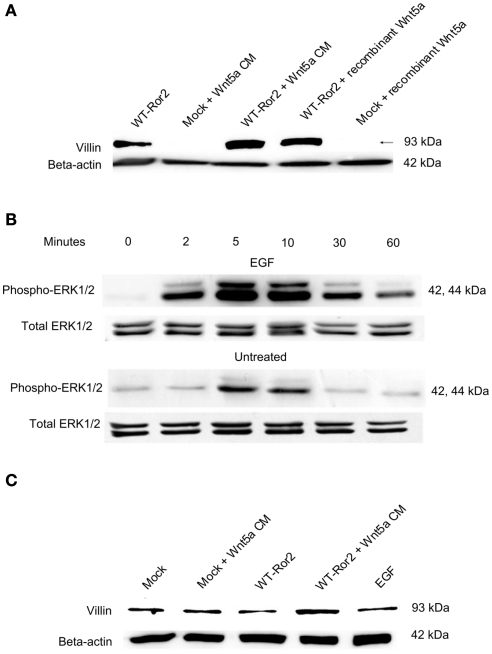
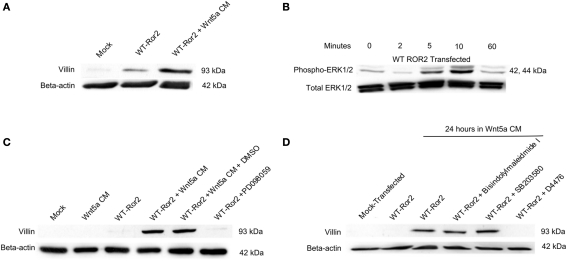
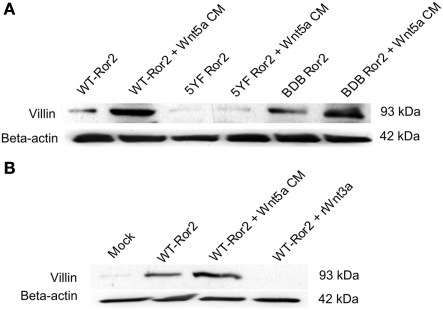
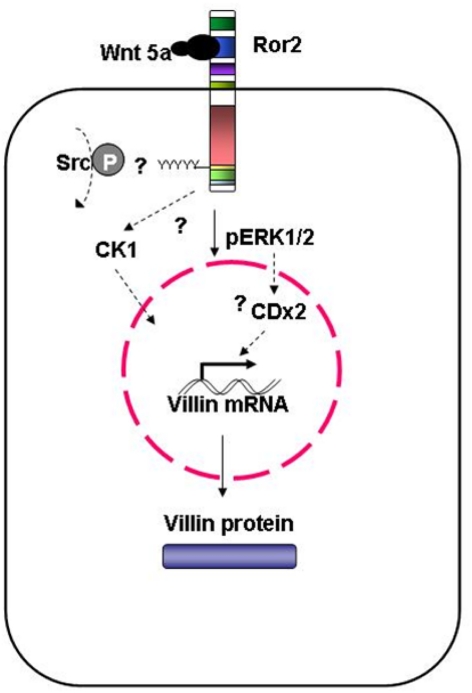
Regulation of expression of the intestinal epithelial actin-binding protein, villin, is poorly understood. The aim of this study was to determine whether Wnt5a stimulates Ror2 in intestinal epithelia caused transient increases in phospho-ERK1/2 (pERK1/2) and subsequently increased expression of villin transcript and protein. To demonstrate Wnt5a-Ror2 regulation of villin expression, we overexpressed wild-type, truncated, or mutant Ror2 constructs in HT29 adenocarcinoma cells and non-transformed fetally derived human intestinal epithelial cells, added conditioned media containing Wnt5a and measured changes in ERK1/2 phosphorylation, villin amplicons, and protein expression by RT-PCR and Western blot techniques. Wnt5a addition caused a transient increase in pERK1/2, which was maximal at 10 min but extinguished by 30 min. Transient transfection with a siRNA duplex against Ror2 diminished Ror2 amplicons and protein and reduced the extent of pERK1/2 activation. Structure-function analysis revealed that the deletion of the cysteine-rich, kringle, or tyrosine kinase domain or substitution mutations of tyrosine residues in the intracellular Ser/Thr-1 region of Ror2 prevented the Wnt5a stimulation of pERK1/2. Deletion of the intracellular proline and serine/threonine-rich regions of Ror2 had no effect on Wnt5a stimulation of pERK1/2. The increase in villin expression was blocked by pharmacological inhibition of MEK-1 and casein kinase 1, but not by PKC and p38 inhibitors. Neither Wnt3a nor epidermal growth factor addition caused increases in villin protein. Our findings suggest that Wnt5a/Ror2 signaling can regulate villin expression in the intestine.
Keywords: Ror2; Wnt signaling; extracellular calcium sensing receptor; villin.
Figures
Similar articles
-
CaSR stimulates secretion of Wnt5a from colonic myofibroblasts to stimulate CDX2 and sucrase-isomaltase using Ror2 on intestinal epithelia.Am J Physiol Gastrointest Liver Physiol. 2008 Oct;295(4):G748-59. doi: 10.1152/ajpgi.00560.2007. Epub 2008 Aug 14. Am J Physiol Gastrointest Liver Physiol. 2008. PMID: 18703641
-
Wnt5a modulates glycogen synthase kinase 3 to induce phosphorylation of receptor tyrosine kinase Ror2.Genes Cells. 2007 Nov;12(11):1215-23. doi: 10.1111/j.1365-2443.2007.01128.x. Genes Cells. 2007. PMID: 17986005
-
Wnt5a secretion stimulated by the extracellular calcium-sensing receptor inhibits defective Wnt signaling in colon cancer cells.Am J Physiol Gastrointest Liver Physiol. 2007 Jul;293(1):G403-11. doi: 10.1152/ajpgi.00119.2007. Epub 2007 Apr 26. Am J Physiol Gastrointest Liver Physiol. 2007. PMID: 17463182
-
Regulation of osteoclast function via Rho-Pkn3-c-Src pathways.J Oral Biosci. 2019 Sep;61(3):135-140. doi: 10.1016/j.job.2019.07.002. Epub 2019 Aug 7. J Oral Biosci. 2019. PMID: 31400545 Review.
-
Insight into the role of Wnt5a-induced signaling in normal and cancer cells.Int Rev Cell Mol Biol. 2015;314:117-48. doi: 10.1016/bs.ircmb.2014.10.003. Epub 2014 Nov 18. Int Rev Cell Mol Biol. 2015. PMID: 25619716 Review.
Cited by
-
Calcium-sensing receptor in colorectal inflammation and cancer: Current insights and future perspectives.World J Gastroenterol. 2018 Sep 28;24(36):4119-4131. doi: 10.3748/wjg.v24.i36.4119. World J Gastroenterol. 2018. PMID: 30271078 Free PMC article. Review.
-
Signaling components of the 1α,25(OH)2D3-dependent Pdia3 receptor complex are required for Wnt5a calcium-dependent signaling.Biochim Biophys Acta. 2014 Nov;1843(11):2365-75. doi: 10.1016/j.bbamcr.2014.06.006. Epub 2014 Jun 16. Biochim Biophys Acta. 2014. PMID: 24946135 Free PMC article.
-
Porcine Intestinal Enteroids: a New Model for Studying Enteric Coronavirus Porcine Epidemic Diarrhea Virus Infection and the Host Innate Response.J Virol. 2019 Feb 19;93(5):e01682-18. doi: 10.1128/JVI.01682-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541861 Free PMC article.
-
Higher expression level of tyrosine kinase-like orphan receptor 2 and Wnt member 5a in papillary thyroid carcinoma is associated with poor prognosis.Oncol Lett. 2017 Nov;14(5):5966-5972. doi: 10.3892/ol.2017.6989. Epub 2017 Sep 18. Oncol Lett. 2017. PMID: 29113233 Free PMC article.
-
Sendai virus recruits cellular villin to remodel actin cytoskeleton during fusion with hepatocytes.Mol Biol Cell. 2017 Dec 15;28(26):3801-3814. doi: 10.1091/mbc.E17-06-0400. Epub 2017 Oct 26. Mol Biol Cell. 2017. PMID: 29074568 Free PMC article.
References
-
- Aliaga J. C., Deschenes C., Beaulieu J. F., Calvo E. L., Rivard N. (1999). Requirement of the MAP kinase cascade for cell cycle progression and differentiation of human intestinal cells. Am. J. Physiol. 277, G631–G641 - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous