

A viral ubiquitin ligase has substrate preferential SUMO targeted ubiquitin ligase activity that counteracts intrinsic antiviral defence
- PMID: 21949651
- PMCID: PMC3174244
- DOI: 10.1371/journal.ppat.1002245
A viral ubiquitin ligase has substrate preferential SUMO targeted ubiquitin ligase activity that counteracts intrinsic antiviral defence
Abstract
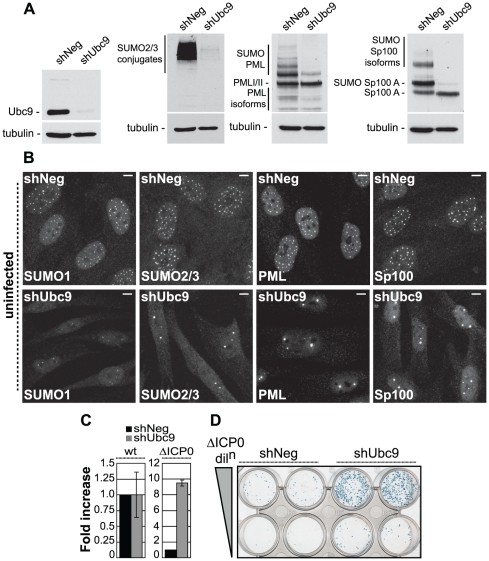
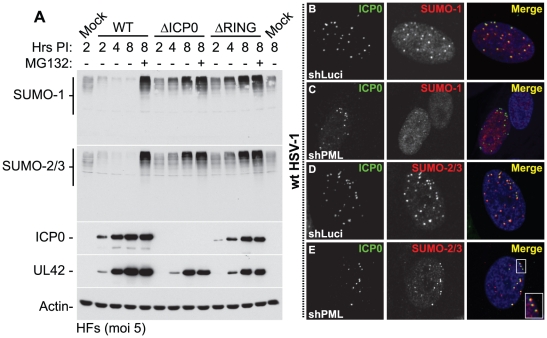
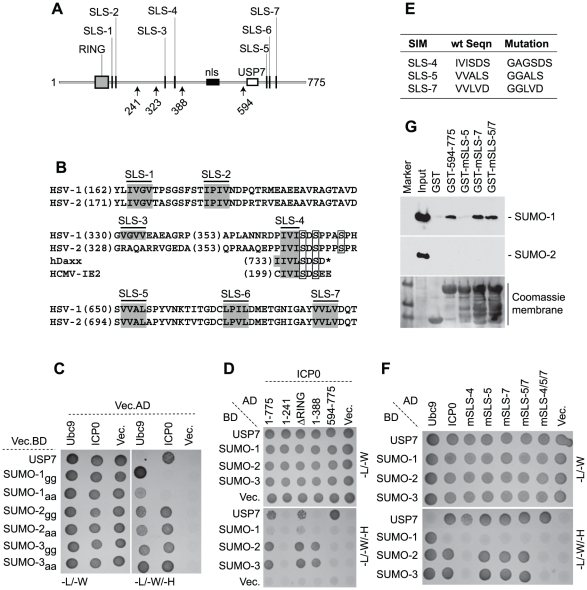
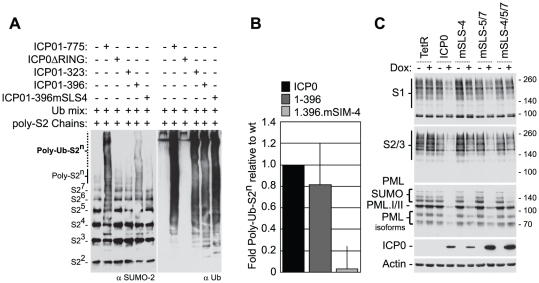
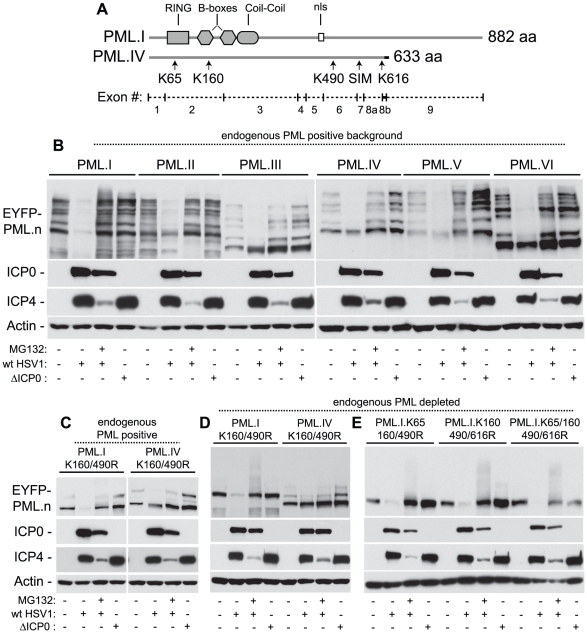
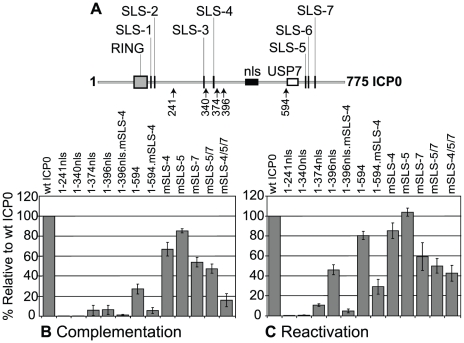
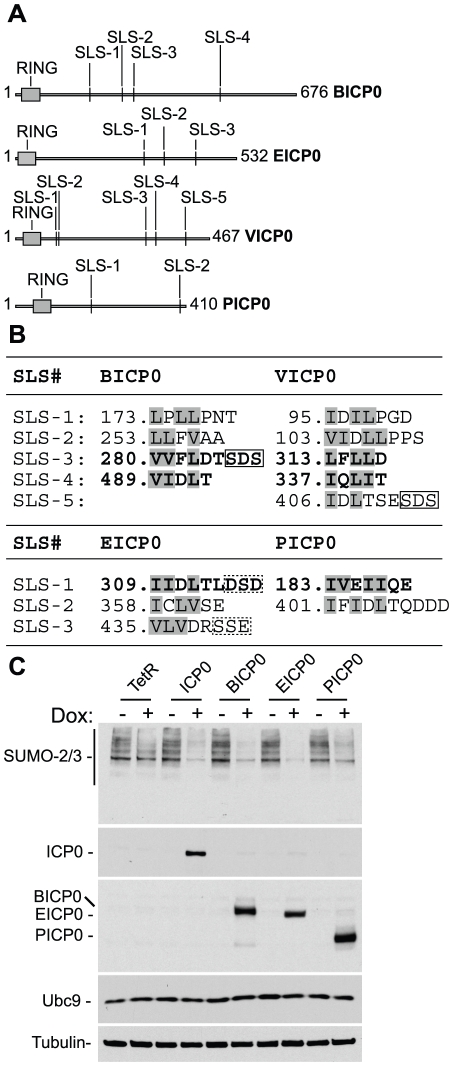
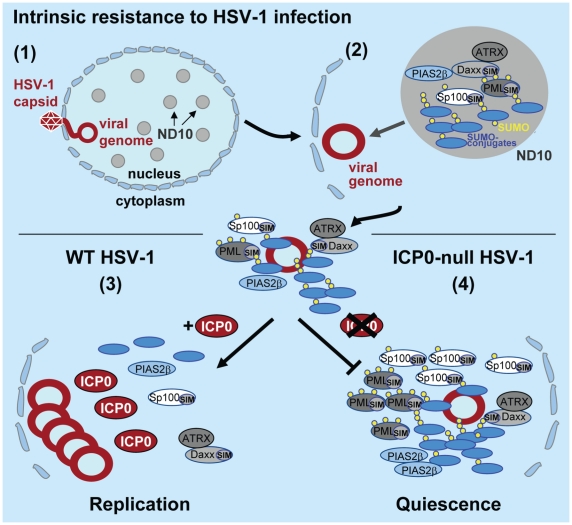
Intrinsic antiviral resistance represents the first line of intracellular defence against virus infection. During herpes simplex virus type-1 (HSV-1) infection this response can lead to the repression of viral gene expression but is counteracted by the viral ubiquitin ligase ICP0. Here we address the mechanisms by which ICP0 overcomes this antiviral response. We report that ICP0 induces the widespread proteasome-dependent degradation of SUMO-conjugated proteins during infection and has properties related to those of cellular SUMO-targeted ubiquitin ligases (STUbLs). Mutation of putative SUMO interaction motifs within ICP0 not only affects its ability to degrade SUMO conjugates, but also its capacity to stimulate HSV-1 lytic infection and reactivation from quiescence. We demonstrate that in the absence of this viral countermeasure the SUMO conjugation pathway plays an important role in mediating intrinsic antiviral resistance and the repression of HSV-1 infection. Using PML as a model substrate, we found that whilst ICP0 preferentially targets SUMO-modified isoforms of PML for degradation, it also induces the degradation of PML isoform I in a SUMO modification-independent manner. PML was degraded by ICP0 more rapidly than the bulk of SUMO-modified proteins in general, implying that the identity of a SUMO-modified protein, as well as the presence of SUMO modification, is involved in ICP0 targeting. We conclude that ICP0 has dual targeting mechanisms involving both SUMO- and substrate-dependent targeting specificities in order to counteract intrinsic antiviral resistance to HSV-1 infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Randow F, Lehner PJ. Viral avoidance and exploitation of the ubiquitin system. Nat Cell Biol. 2009;11:527–534. - PubMed
-
- Everett RD. The roles of ICP0 during HSV-1 infection. Alpha herpesviruses: molecular and cellular biology. In: Sandri-Goldin RM, editor. Wymondham, United Kingdom: Caister Academic Press; 2006b. pp. 39–64.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
