

Secreted protein acidic and rich in cysteine is a matrix scavenger chaperone
- PMID: 21949685
- PMCID: PMC3174944
- DOI: 10.1371/journal.pone.0023880
Secreted protein acidic and rich in cysteine is a matrix scavenger chaperone
Abstract
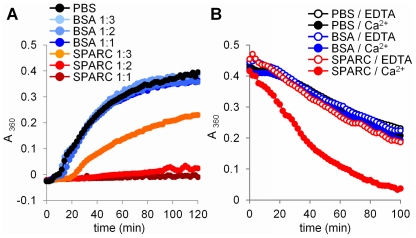
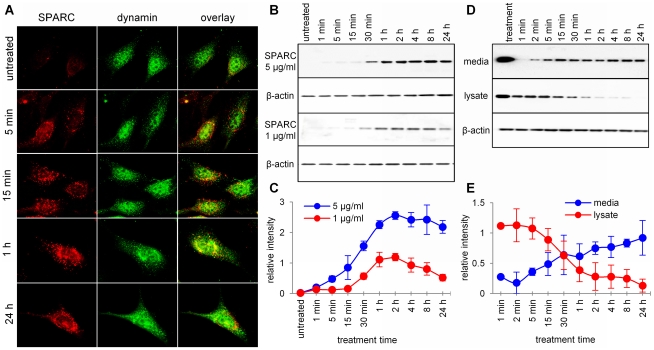
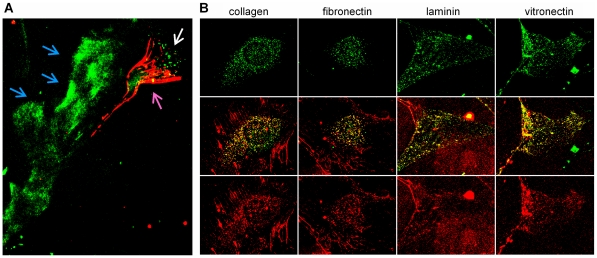
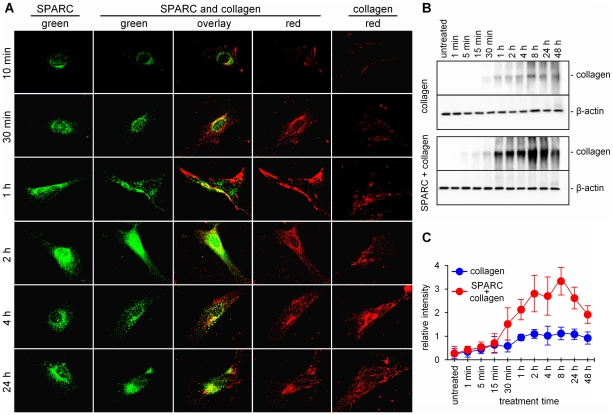
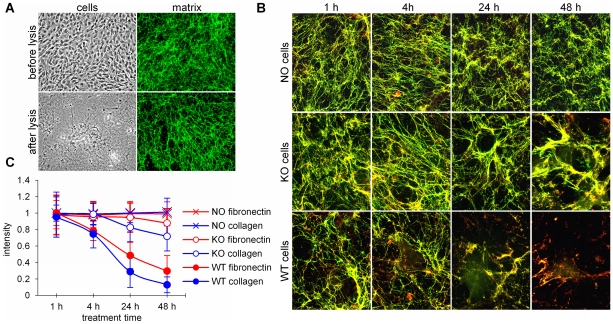
Secreted Protein Acidic and Rich in Cysteine (SPARC) is one of the major non-structural proteins of the extracellular matrix (ECM) in remodeling tissues. The functional significance of SPARC is emphasized by its origin in the first multicellular organisms and its high degree of evolutionary conservation. Although SPARC has been shown to act as a critical modulator of ECM remodeling with profound effects on tissue physiology and architecture, no plausible molecular mechanism of its action has been proposed. In the present study, we demonstrate that SPARC mediates the disassembly and degradation of ECM networks by functioning as a matricellular chaperone. While it has low affinity to its targets inside the cells where the Ca(2+) concentrations are low, high extracellular concentrations of Ca(2+) activate binding to multiple ECM proteins, including collagens. We demonstrated that in vitro, this leads to the inhibition of collagen I fibrillogenesis and disassembly of pre-formed collagen I fibrils by SPARC at high Ca(2+) concentrations. In cell culture, exogenous SPARC was internalized by the fibroblast cells in a time- and concentration-dependent manner. Pulse-chase assay further revealed that internalized SPARC is quickly released outside the cell, demonstrating that SPARC shuttles between the cell and ECM. Fluorescently labeled collagen I, fibronectin, vitronectin, and laminin were co-internalized with SPARC by fibroblasts, and semi-quantitative Western blot showed that SPARC mediates internalization of collagen I. Using a novel 3-dimensional model of fluorescent ECM networks pre-deposited by live fibroblasts, we demonstrated that degradation of ECM depends on the chaperone activity of SPARC. These results indicate that SPARC may represent a new class of scavenger chaperones, which mediate ECM degradation, remodeling and repair by disassembling ECM networks and shuttling ECM proteins into the cell. Further understanding of this mechanism may provide insight into the pathogenesis of matrix-associated disorders and lead to the novel treatment strategies.
Conflict of interest statement
Figures
Similar articles
-
Type I collagen-deficient Mov-13 mice do not retain SPARC in the extracellular matrix: implications for fibroblast function.Dev Dyn. 1996 Oct;207(2):171-83. doi: 10.1002/(SICI)1097-0177(199610)207:2<171::AID-AJA5>3.0.CO;2-E. Dev Dyn. 1996. PMID: 8906420
-
Cardiac extracellular matrix remodeling: fibrillar collagens and Secreted Protein Acidic and Rich in Cysteine (SPARC).J Mol Cell Cardiol. 2010 Mar;48(3):544-9. doi: 10.1016/j.yjmcc.2009.06.018. Epub 2009 Jul 3. J Mol Cell Cardiol. 2010. PMID: 19577572 Free PMC article. Review.
-
Effect of Sparc knockout on the extracellular matrix of mouse periodontal ligament cells.Biochem Biophys Res Commun. 2024 Jan 15;692:149364. doi: 10.1016/j.bbrc.2023.149364. Epub 2023 Dec 6. Biochem Biophys Res Commun. 2024. PMID: 38070276
-
The Influence of the Extracellular Matrix in Inflammation: Findings from the SPARC-Null Mouse.Anat Rec (Hoboken). 2020 Jun;303(6):1624-1629. doi: 10.1002/ar.24133. Epub 2019 Apr 24. Anat Rec (Hoboken). 2020. PMID: 30980479 Free PMC article. Review.
-
A 9-kDa matricellular SPARC fragment released by cathepsin D exhibits pro-tumor activity in the triple-negative breast cancer microenvironment.Theranostics. 2021 Apr 15;11(13):6173-6192. doi: 10.7150/thno.58254. eCollection 2021. Theranostics. 2021. PMID: 33995652 Free PMC article.
Cited by
-
Increased Rate of Retinal Pigment Epithelial Cell Migration and Pro-Angiogenic Potential Ensuing From Reduced Cystatin C Expression.Invest Ophthalmol Vis Sci. 2020 Feb 7;61(2):9. doi: 10.1167/iovs.61.2.9. Invest Ophthalmol Vis Sci. 2020. PMID: 32049341 Free PMC article.
-
Dynamic regulation of basement membrane protein levels promotes egg chamber elongation in Drosophila.Dev Biol. 2015 Oct 15;406(2):212-21. doi: 10.1016/j.ydbio.2015.08.018. Epub 2015 Sep 6. Dev Biol. 2015. PMID: 26348027 Free PMC article.
-
Identifying Potential Candidate Hub Genes and Functionally Enriched Pathways in the Immune Responses to Quadrivalent Inactivated Influenza Vaccines in the Elderly Through Co-Expression Network Analysis.Front Immunol. 2020 Dec 4;11:603337. doi: 10.3389/fimmu.2020.603337. eCollection 2020. Front Immunol. 2020. PMID: 33343577 Free PMC article. Clinical Trial.
-
Thrombospondin expression in myofibers stabilizes muscle membranes.Elife. 2016 Sep 26;5:e17589. doi: 10.7554/eLife.17589. Elife. 2016. PMID: 27669143 Free PMC article.
-
Overexpression of SPARC in human trabecular meshwork increases intraocular pressure and alters extracellular matrix.Invest Ophthalmol Vis Sci. 2013 May 7;54(5):3309-19. doi: 10.1167/iovs.12-11362. Invest Ophthalmol Vis Sci. 2013. PMID: 23599341 Free PMC article.
References
-
- Chlenski A, Cohn SL. Modulation of matrix remodeling by SPARC in neoplastic progression. Semin Cell Dev Biol. 2010;21:55–65. - PubMed
-
- Kaufmann B, Muller S, Hanisch FG, Hartmann U, Paulsson M, et al. Structural variability of BM-40/SPARC/osteonectin glycosylation: implications for collagen affinity. Glycobiology. 2004;14:609–619. - PubMed
-
- Chlenski A, Liu S, Baker LJ, Yang Q, Tian Y, et al. Neuroblastoma angiogenesis is inhibited with a folded synthetic molecule corresponding to the epidermal growth factor-like module of the follistatin domain of SPARC. Cancer Res. 2004;64:7420–7425. - PubMed
-
- Sasaki T, Gohring W, Mann K, Maurer P, Hohenester E, et al. Limited cleavage of extracellular matrix protein BM-40 by matrix metalloproteinases increases its affinity for collagens. J Biol Chem. 1997;272:9237–9243. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
