

Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis
- PMID: 21951689
- PMCID: PMC3193812
- DOI: 10.1186/1741-7007-9-64
Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis
Abstract
Background: The mitochondrial genome of higher plants is unusually dynamic, with recombination and nonhomologous end-joining (NHEJ) activities producing variability in size and organization. Plant mitochondrial DNA also generally displays much lower nucleotide substitution rates than mammalian or yeast systems. Arabidopsis displays these features and expedites characterization of the mitochondrial recombination surveillance gene MSH1 (MutS 1 homolog), lending itself to detailed study of de novo mitochondrial genome activity. In the present study, we investigated the underlying basis for unusual plant features as they contribute to rapid mitochondrial genome evolution.
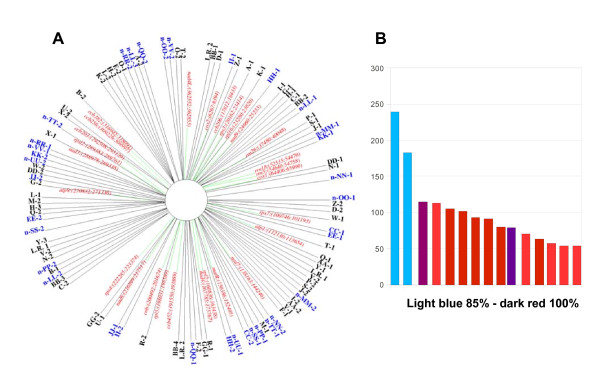
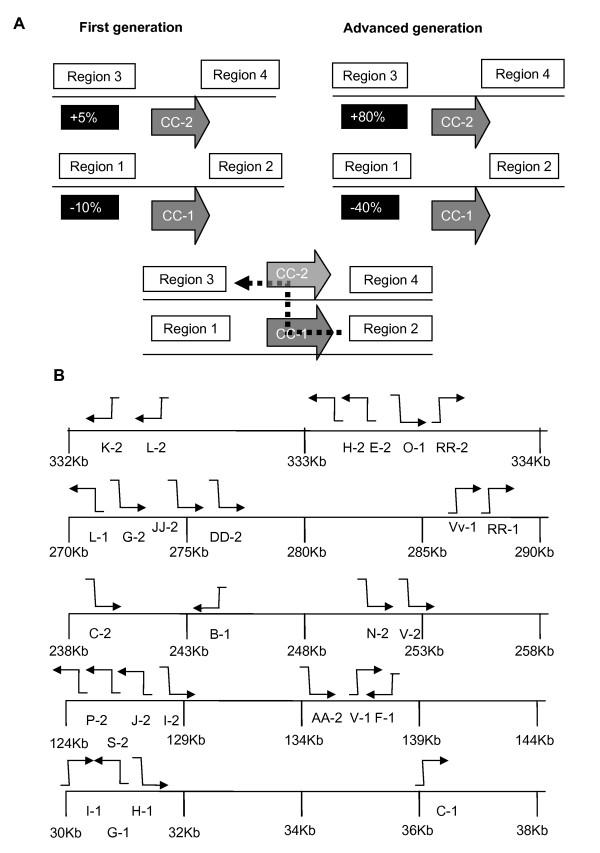
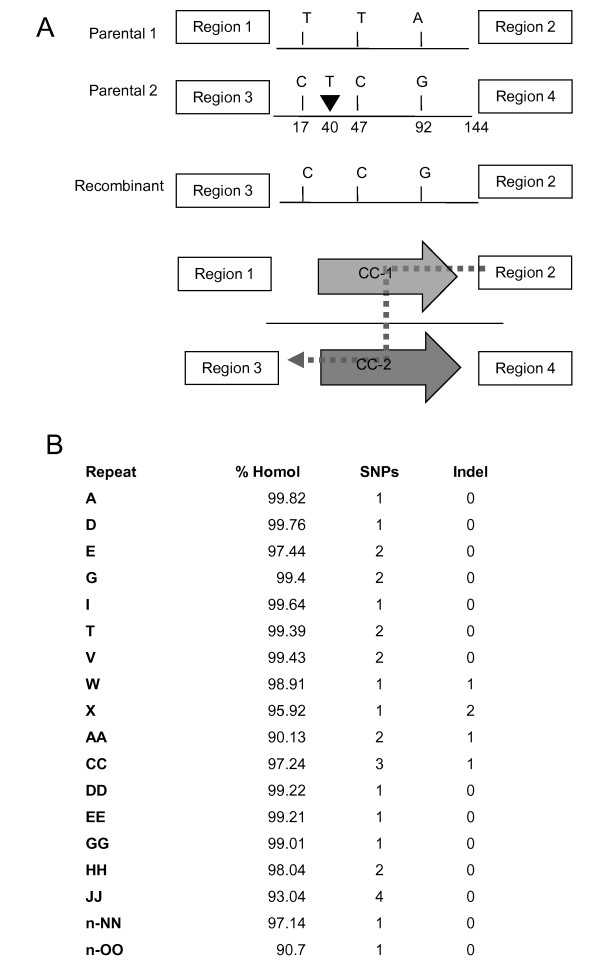
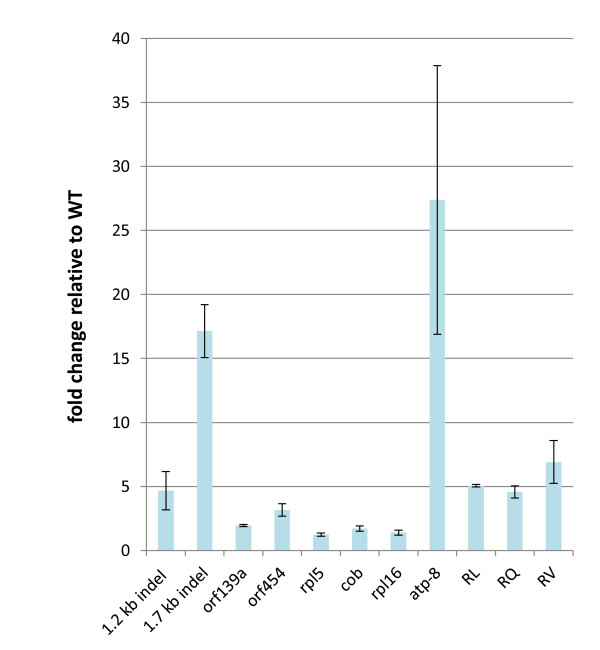
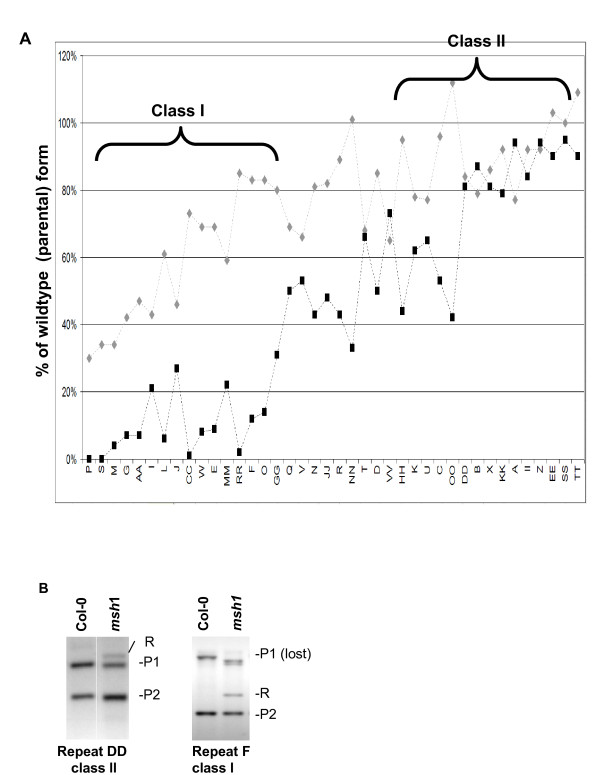
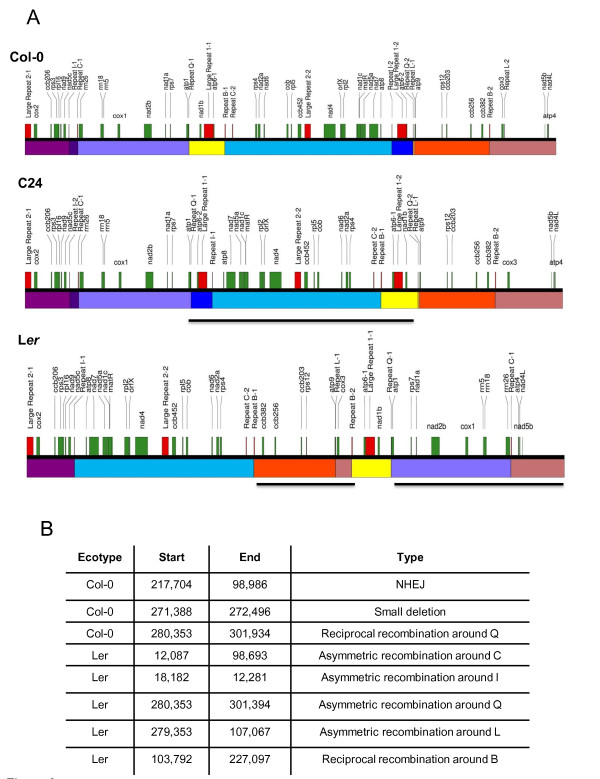
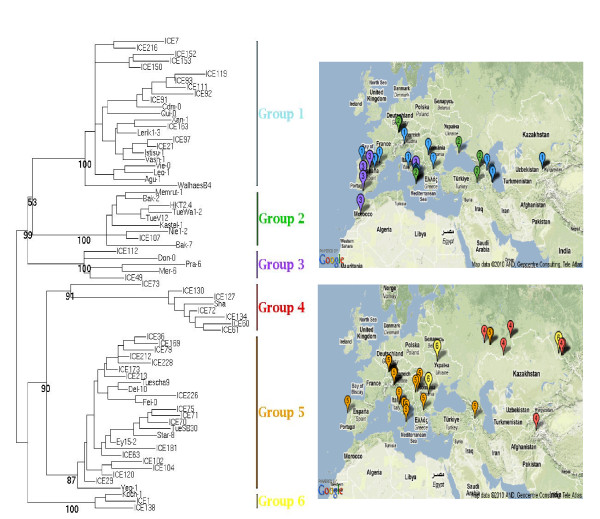
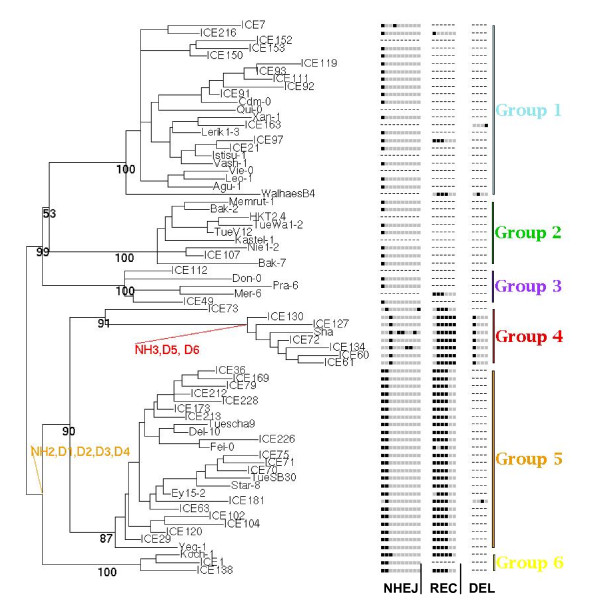
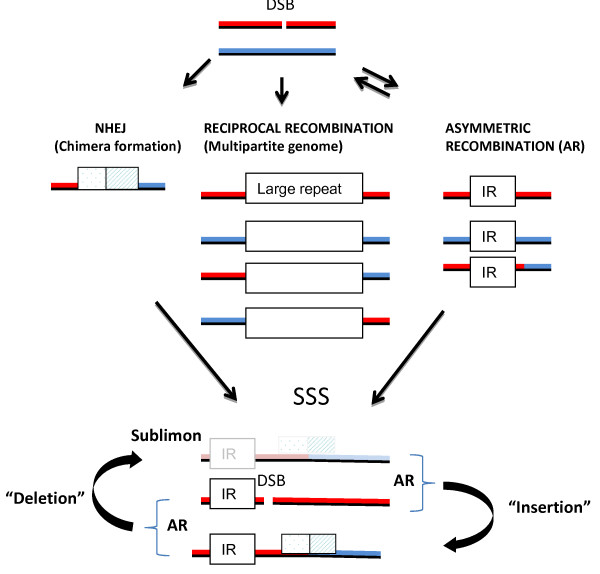
Results: We obtained evidence of double-strand break (DSB) repair, including NHEJ, sequence deletions and mitochondrial asymmetric recombination activity in Arabidopsis wild-type and msh1 mutants on the basis of data generated by Illumina deep sequencing and confirmed by DNA gel blot analysis. On a larger scale, with mitochondrial comparisons across 72 Arabidopsis ecotypes, similar evidence of DSB repair activity differentiated ecotypes. Forty-seven repeat pairs were active in DNA exchange in the msh1 mutant. Recombination sites showed asymmetrical DNA exchange within lengths of 50- to 556-bp sharing sequence identity as low as 85%. De novo asymmetrical recombination involved heteroduplex formation, gene conversion and mismatch repair activities. Substoichiometric shifting by asymmetrical exchange created the appearance of rapid sequence gain and loss in association with particular repeat classes.
Conclusions: Extensive mitochondrial genomic variation within a single plant species derives largely from DSB activity and its repair. Observed gene conversion and mismatch repair activity contribute to the low nucleotide substitution rates seen in these genomes. On a phenotypic level, these patterns of rearrangement likely contribute to the reproductive versatility of higher plants.
Figures
Comment in
-
The intriguing evolutionary dynamics of plant mitochondrial DNA.BMC Biol. 2011 Sep 27;9:61. doi: 10.1186/1741-7007-9-61. BMC Biol. 2011. PMID: 21951676 Free PMC article.
References
-
- Sugiyama Y, Watase Y, Nagase M, Makita N, Yagura S, Hirai A, Sugiura M. The complete nucleotide sequence and multipartite organization of the tobacco mitochondrial genome: comparative analysis of mitochondrial genomes in higher plants. Mol Gen Genomics. 2005;272:603–615. doi: 10.1007/s00438-004-1075-8. - DOI - PubMed
-
- Arrieta-Montiel MP, Mackenzie SA. In: Advances in Plant Biology: Plant Mitochondria. Kempen F, editor. New York: Springer; 2010. Plant mitochondrial genomes and recombination; pp. 65–84.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
