

The MHC I immunopeptidome conveys to the cell surface an integrative view of cellular regulation
- PMID: 21952136
- PMCID: PMC3202804
- DOI: 10.1038/msb.2011.68
The MHC I immunopeptidome conveys to the cell surface an integrative view of cellular regulation
Abstract
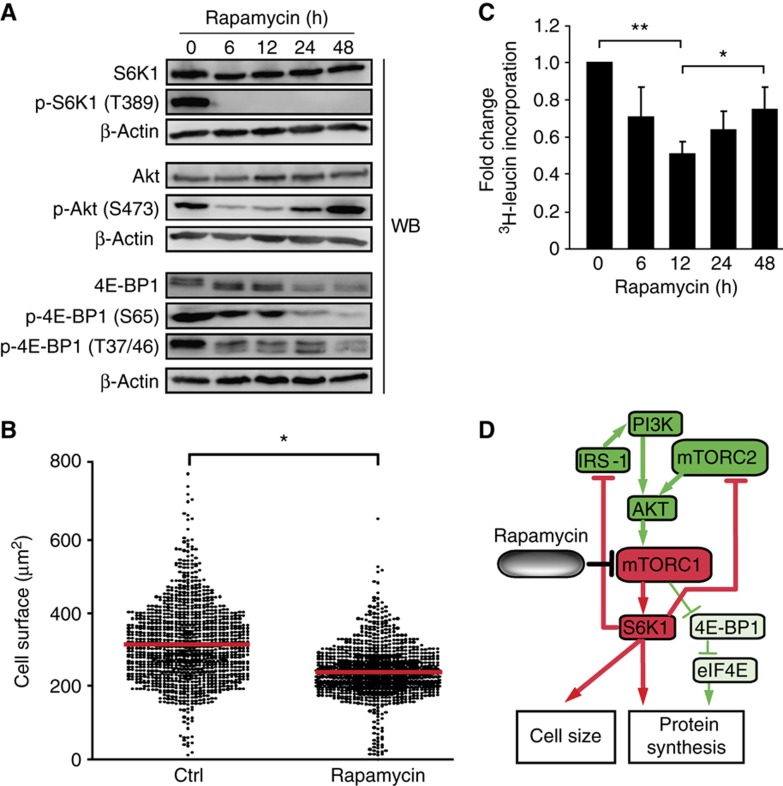
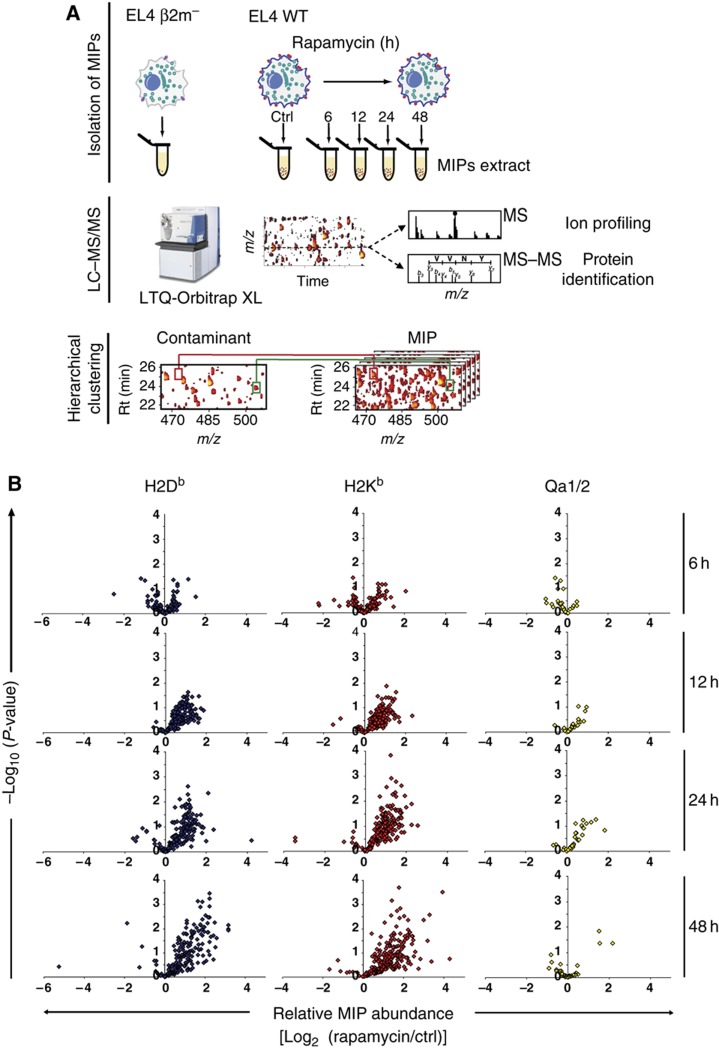
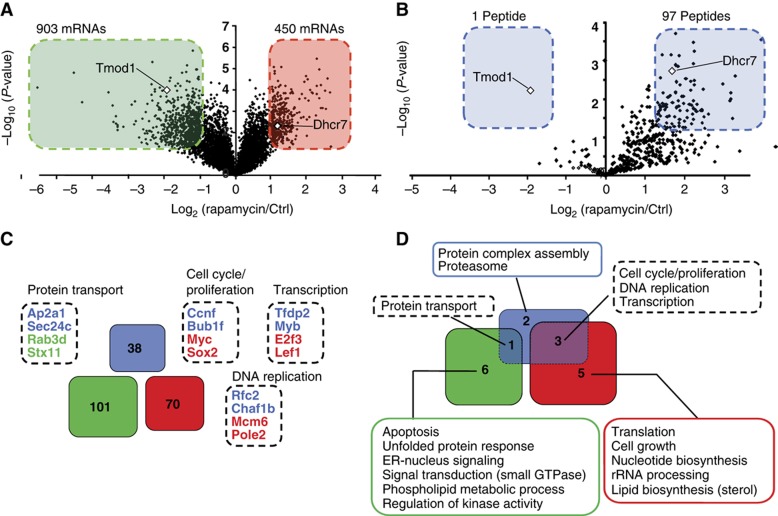
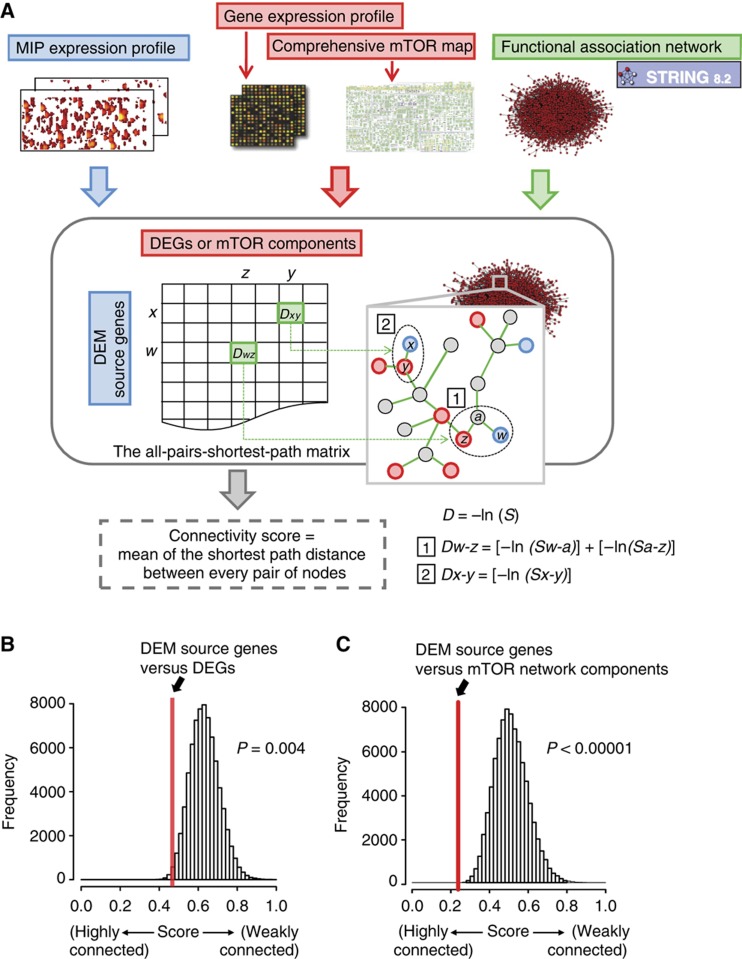
Self/non-self discrimination is a fundamental requirement of life. Endogenous peptides presented by major histocompatibility complex class I (MHC I) molecules represent the essence of self for CD8 T lymphocytes. These MHC I peptides (MIPs) are collectively referred to as the immunopeptidome. From a systems-level perspective, very little is known about the origin, composition and plasticity of the immunopeptidome. Here, we show that the immunopeptidome, and therefore the nature of the immune self, is plastic and moulded by cellular metabolic activity. By using a quantitative high-throughput mass spectrometry-based approach, we found that altering cellular metabolism via the inhibition of the mammalian target of rapamycin results in dynamic changes in the cell surface MIPs landscape. Moreover, we provide systems-level evidence that the immunopeptidome projects at the cell surface a representation of biochemical networks and metabolic events regulated at multiple levels inside the cell. Our findings open up new perspectives in systems immunology and predictive biology. Indeed, predicting variations in the immunopeptidome in response to cell-intrinsic and -extrinsic factors could be relevant to the rational design of immunotherapeutic interventions.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
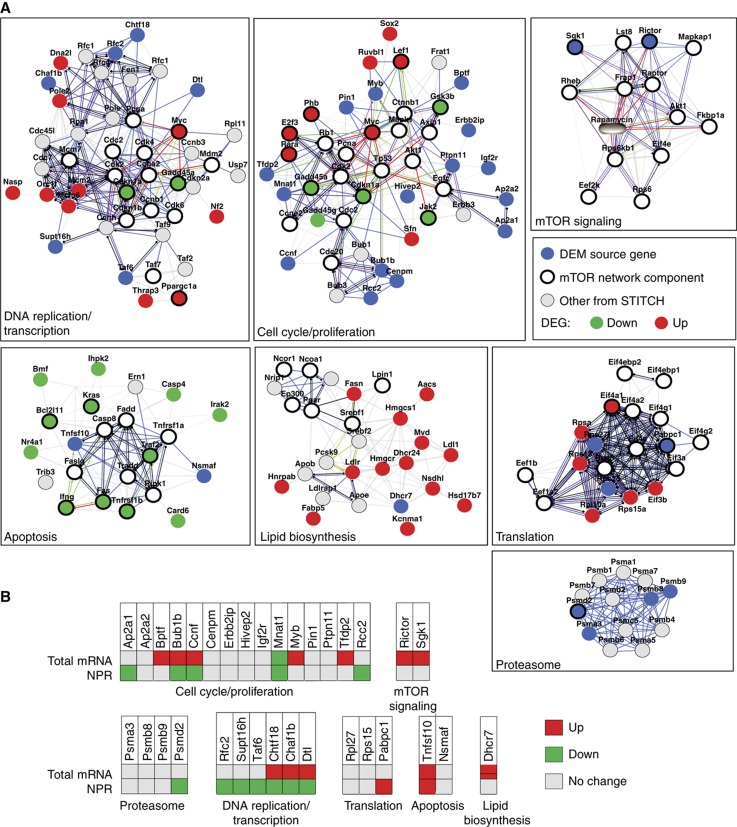
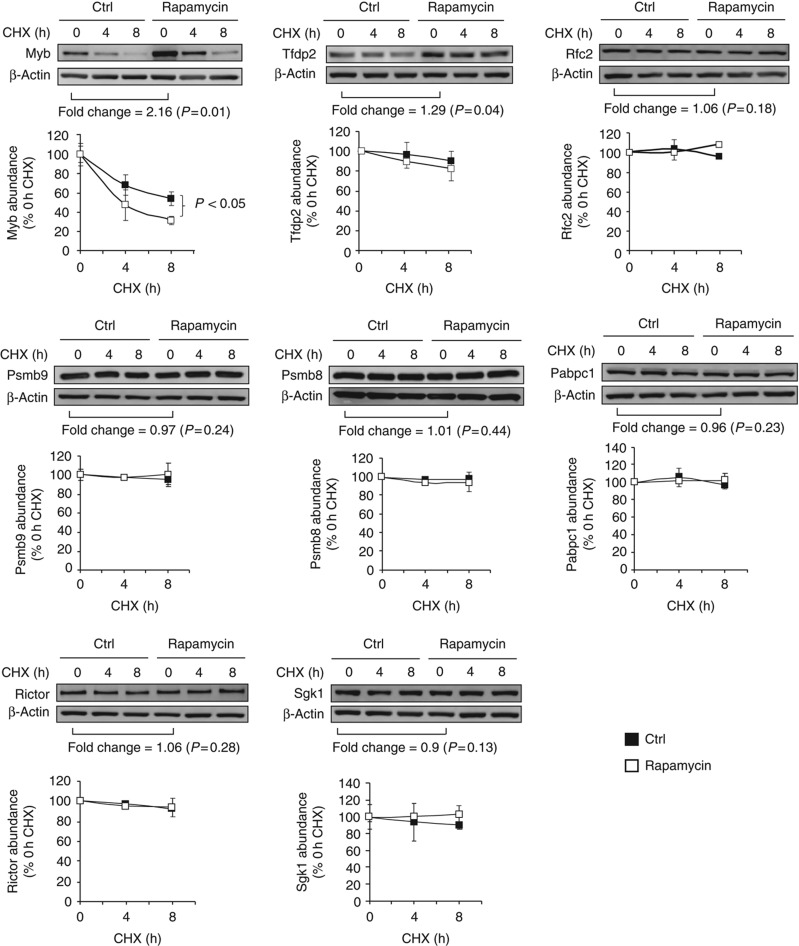
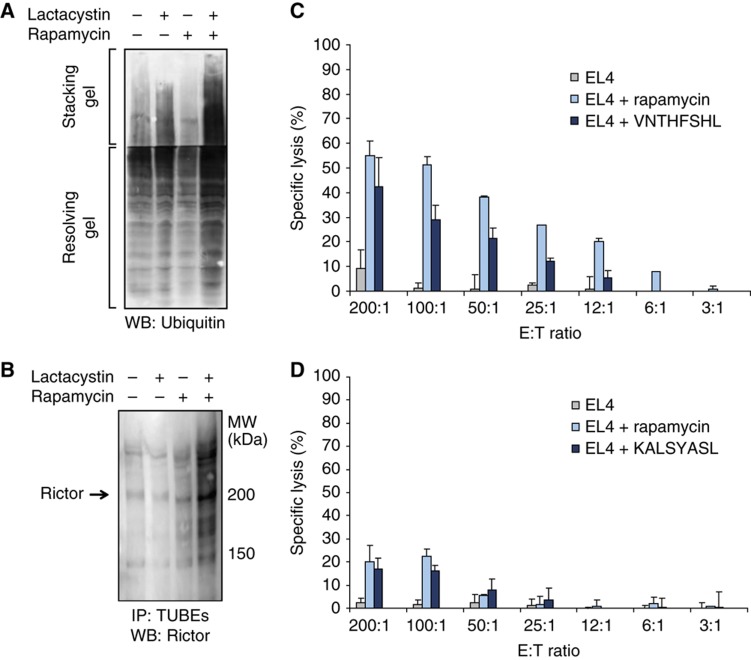
References
-
- Barnea E, Beer I, Patoka R, Ziv T, Kessler O, Tzehoval E, Eisenbach L, Zavazava N, Admon A (2002) Analysis of endogenous peptides bound by soluble MHC class I molecules: a novel approach for identifying tumor-specific antigens. Eur J Immunol 32: 213–222 - PubMed
-
- Benoist C, Germain RN, Mathis D (2006) A plaidoyer for ‘systems immunology’. Immunol Rev 210: 229–234 - PubMed
-
- Boehm T (2006) Quality control in self/nonself discrimination. Cell 125: 845–858 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
