

From revolution to evolution: the glutamate hypothesis of schizophrenia and its implication for treatment
- PMID: 21956446
- PMCID: PMC3238069
- DOI: 10.1038/npp.2011.181
From revolution to evolution: the glutamate hypothesis of schizophrenia and its implication for treatment
Abstract
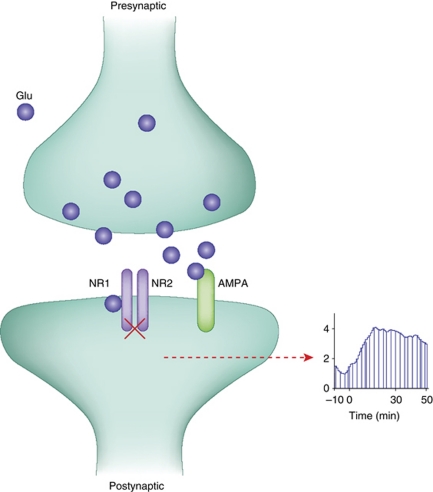
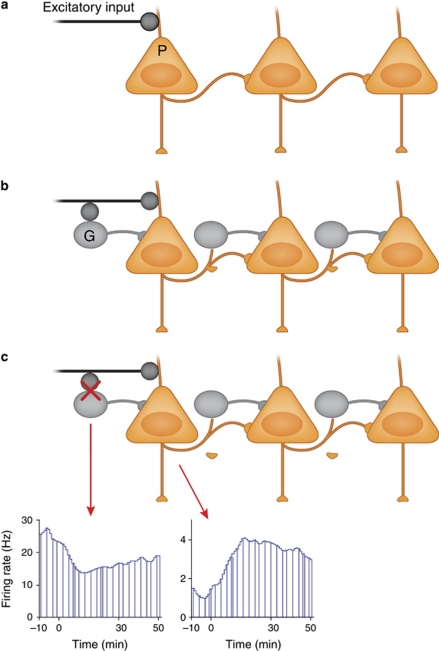
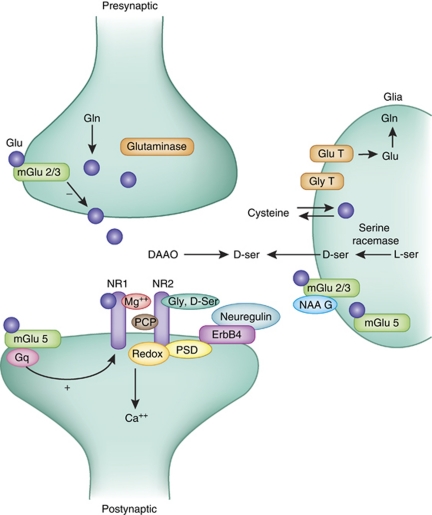
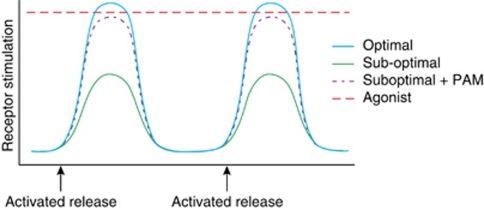
Glutamate is the primary excitatory neurotransmitter in mammalian brain. Disturbances in glutamate-mediated neurotransmission have been increasingly documented in a range of neuropsychiatric disorders including schizophrenia, substance abuse, mood disorders, Alzheimer's disease, and autism-spectrum disorders. Glutamatergic theories of schizophrenia are based on the ability of N-methyl-D-aspartate receptor (NMDAR) antagonists to induce schizophrenia-like symptoms, as well as emergent literature documenting disturbances of NMDAR-related gene expression and metabolic pathways in schizophrenia. Research over the past two decades has highlighted promising new targets for drug development based on potential pre- and postsynaptic, and glial mechanisms leading to NMDAR dysfunction. Reduced NMDAR activity on inhibitory neurons leads to disinhibition of glutamate neurons increasing synaptic activity of glutamate, especially in the prefrontal cortex. Based on this mechanism, normalizing excess glutamate levels by metabotropic glutamate group 2/3 receptor agonists has led to potential identification of the first non-monoaminergic target with comparable efficacy as conventional antipsychotic drugs for treating positive and negative symptoms of schizophrenia. In addition, NMDAR has intrinsic modulatory sites that are active targets for drug development, several of which show promise in preclinical/early clinical trials targeting both symptoms and cognition. To date, most studies have been done with orthosteric agonists and/or antagonists at specific sites. However, allosteric modulators, both positive and negative, may offer superior efficacy with less danger of downregulation.
Figures
References
-
- Abbott A. Schizophrenia: the drug deadlock. Nature. 2010;468:158–159. - PubMed
-
- Allen NC, Bagade S, McQueen MB, Ioannidis JP, Kavvoura FK, Khoury MJ, et al. Systematic meta-analyses and field synopsis of genetic association studies in schizophrenia: the SzGene database. Nat Genet. 2008;40:827–834. - PubMed
-
- Anderson S, Volk D, et al. Increased density of microtubule associated protein 2-immunoreactive neurons in the prefrontal white matter of schizophrenic subjects. Schizophr Res. 1996;19:111–119. - PubMed
-
- Arai AC, Kessler M. Pharmacology of ampakine modulators: from AMPA receptors to synapses and behavior. Curr Drug Targets. 2007;8:583–602. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
