

Viruses and the cellular RNA decay machinery
- PMID: 21956906
- PMCID: PMC7169783
- DOI: 10.1002/wrna.3
Viruses and the cellular RNA decay machinery
Abstract
The ability to control cellular and viral gene expression, either globally or selectively, is central to a successful viral infection, and it is also crucial for the host to respond and eradicate pathogens. In eukaryotes, regulation of message stability contributes significantly to the control of gene expression and plays a prominent role in the normal physiology of a cell as well as in its response to environmental and pathogenic stresses. Not surprisingly, emerging evidence indicates that there are significant interactions between the eukaryotic RNA turnover machinery and a wide variety of viruses. Interestingly, in many cases viruses have evolved mechanisms not only to evade eradication by these pathways, but also to manipulate them for enhanced viral replication and gene expression. Given our incomplete understanding of how many of these pathways are normally regulated, viruses should be powerful tools to help deconstruct the complex networks and events governing eukaryotic RNA stability.
2010 John Wiley & Sons, Ltd.
Figures
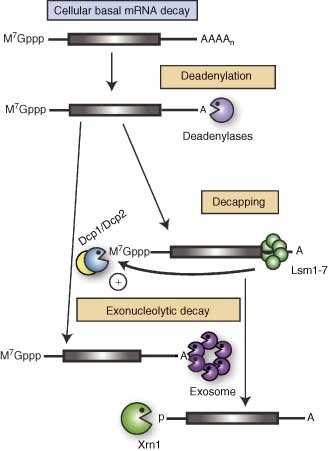
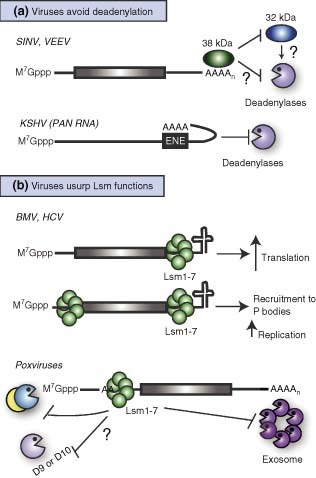
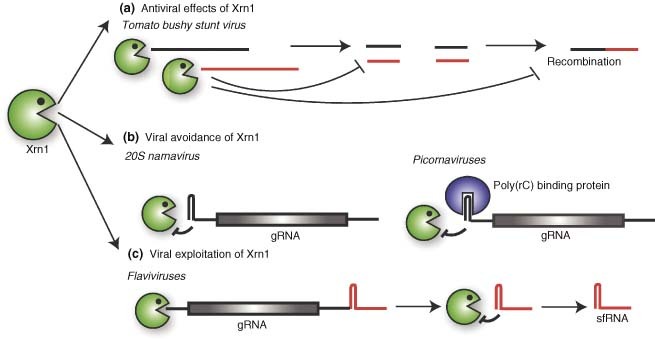
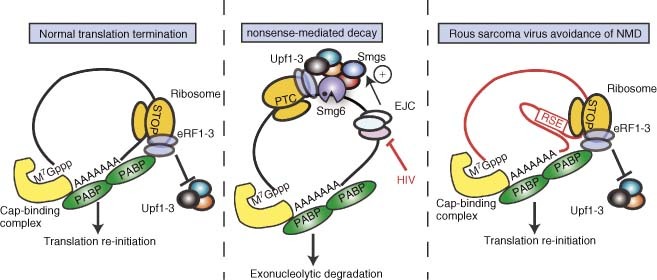
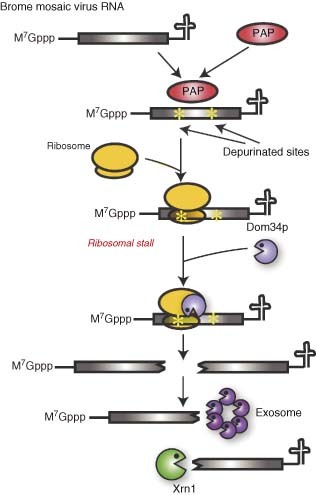
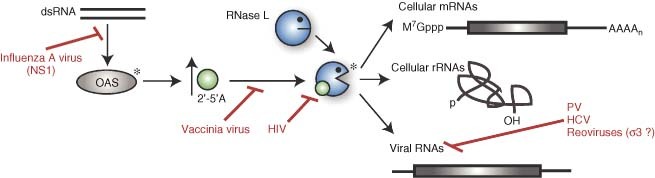
References
-
- Doma MK, Parker R. RNA quality control in eukaryotes. Cell 2007, 131: 660–668. - PubMed
-
- Goldstrohm AC, Wickens M. Multifunctional deadenylase complexes diversify mRNA control. Nat Rev Mol Cell Biol 2008, 9: 337–344. - PubMed
-
- Parker R, Song H. The enzymes and control of eukaryotic mRNA turnover. Nat Struct Mol Biol 2004, 11: 121–127. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
