

Measurements of the acidification kinetics of single SynaptopHluorin vesicles
- PMID: 21961583
- PMCID: PMC3183813
- DOI: 10.1016/j.bpj.2011.08.032
Measurements of the acidification kinetics of single SynaptopHluorin vesicles
Abstract
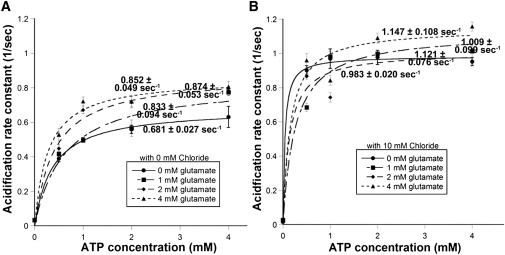
Uptake of neurotransmitters into synaptic vesicles is driven by the proton gradient established across the vesicle membrane. The acidification of synaptic vesicles, therefore, is a crucial component of vesicle function. Here we present measurements of acidification rate constants from isolated, single synaptic vesicles. Vesicles were purified from mice expressing a fusion protein termed SynaptopHluorin created by the fusion of VAMP/synaptobrevin to the pH-sensitive super-ecliptic green fluorescent protein. We calibrated SynaptopHluorin fluorescence to determine the relationship between fluorescence intensity and internal vesicle pH, and used these values to measure the rate constant of vesicle acidification. We also measured the effects of ATP, glutamate, and chloride on acidification. We report acidification time constants of 500 ms to 1 s. The rate of acidification increased with increasing extravesicular concentrations of ATP and glutamate. These data provide an upper and a lower bound for vesicle acidification and indicate that vesicle readiness can be regulated by changes in energy and transmitter availability.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
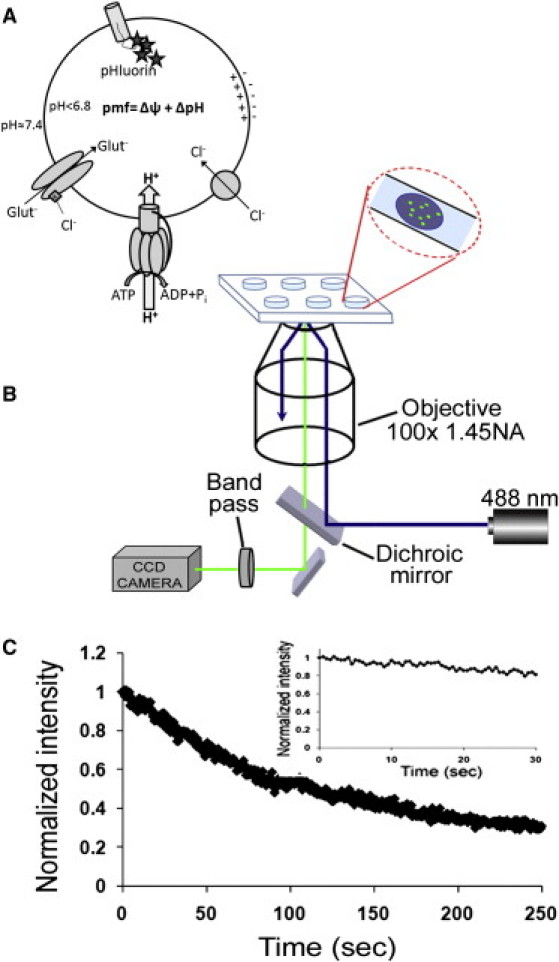
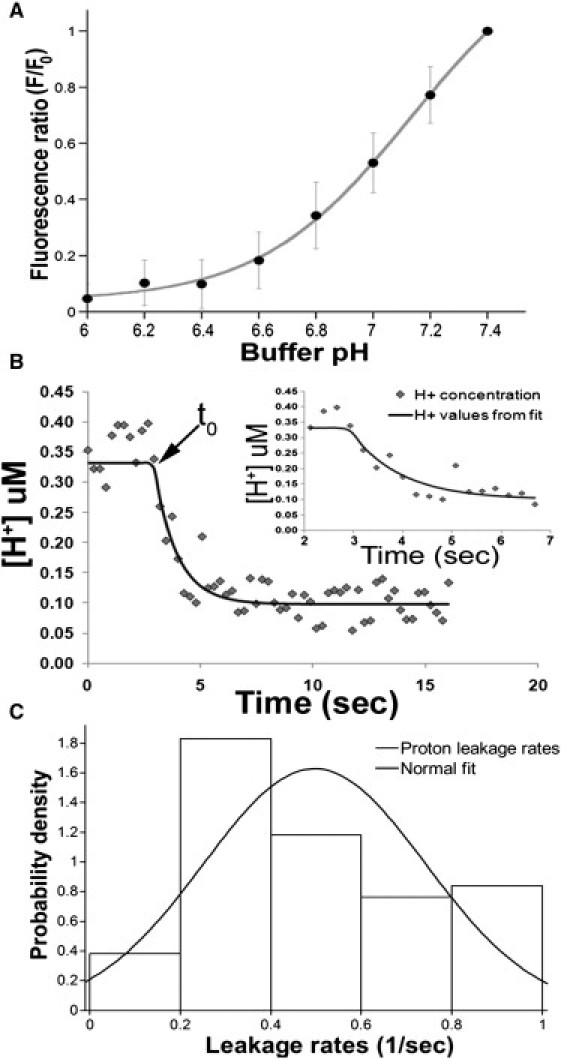
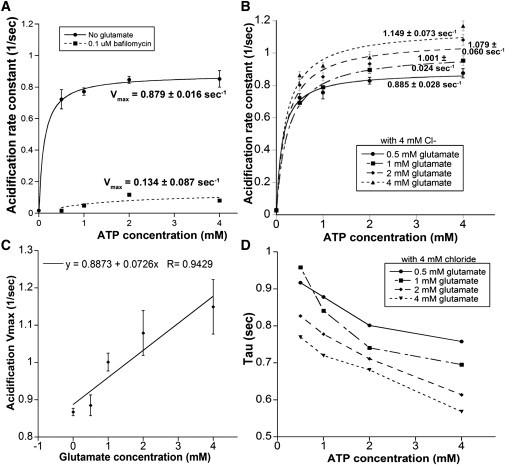
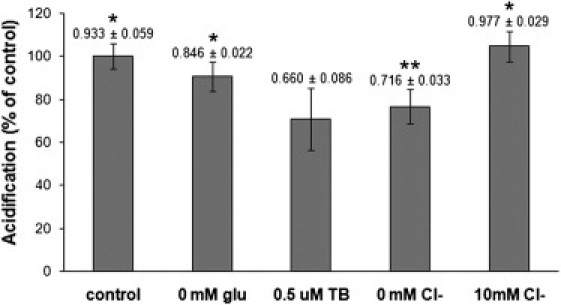
References
-
- Naito S., Ueda T. Characterization of glutamate uptake into synaptic vesicles. J. Neurochem. 1985;44:99–109. - PubMed
-
- Cidon S., Sihra T.S. Characterization of a H+-ATPase in rat brain synaptic vesicles. Coupling to L-glutamate transport. J. Biol. Chem. 1989;264:8281–8288. - PubMed
-
- Wolosker H., de Souza D.O., de Meis L. Regulation of glutamate transport into synaptic vesicles by chloride and proton gradient. J. Biol. Chem. 1996;271:11726–11731. - PubMed
-
- Clerc S., Barenholz Y. A quantitative model for using acridine orange as a transmembrane pH gradient probe. Anal. Biochem. 1998;259:104–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
