

Nuclear actin-related protein is required for chromosome segregation in Toxoplasma gondii
- PMID: 21963440
- PMCID: PMC3767130
- DOI: 10.1016/j.molbiopara.2011.09.006
Nuclear actin-related protein is required for chromosome segregation in Toxoplasma gondii
Abstract
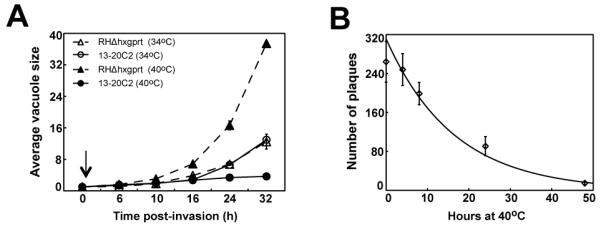
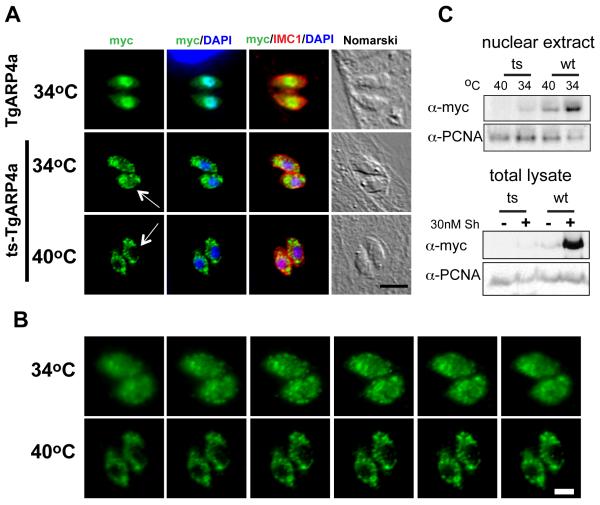
Apicomplexa parasites use complex cell cycles to replicate that are not well understood mechanistically. We have established a robust forward genetic strategy to identify the essential components of parasite cell division. Here we describe a novel temperature sensitive Toxoplasma strain, mutant 13-20C2, which growth arrests due to a defect in mitosis. The primary phenotype is the mis-segregation of duplicated chromosomes with chromosome loss during nuclear division. This defect is conditional-lethal with respect to temperature, although relatively mild in regard to the preservation of the major microtubule organizing centers. Despite severe DNA loss many of the physical structures associated with daughter budding and the assembly of invasion structures formed and operated normally at the non-permissive temperature before completely arresting. These results suggest there are coordinating mechanisms that govern the timing of these events in the parasite cell cycle. The defect in mutant 13-20C2 was mapped by genetic complementation to Toxoplasma chromosome III and to a specific mutation in the gene encoding an ortholog of nuclear actin-related protein 4. A change in a conserved isoleucine to threonine in the helical structure of this nuclear actin related protein leads to protein instability and cellular mis-localization at the higher temperature. Given the age of this protist family, the results indicate a key role for nuclear actin-related proteins in chromosome segregation was established very early in the evolution of eukaryotes.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures



References
-
- Dubey JP, Carpenter JL. Toxoplasma gondii-like schizonts in the tracheal epithelium of a cat. J Parasitol. 1991;77:792–6. - PubMed
-
- Canning EU, Sinden RE. The organization of the ookinete and observations on nuclear division in oocysts of Plasmodium berghei. Parasitology. 1973;67:29–40. - PubMed
-
- Speer CA, Dubey JP. An ultrastructural study of first- and second-generation merogony in the coccidian Sarcocystis tenella. J Protozool. 1981;28:424–31. - PubMed
-
- Sheffield HG, Melton ML. The fine structure and reproduction of Toxoplasma gondii. J Parasitol. 1968;54:209–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
