

Functional dissection of phosphorylation of Disheveled in Drosophila
- PMID: 21963539
- PMCID: PMC3221411
- DOI: 10.1016/j.ydbio.2011.09.017
Functional dissection of phosphorylation of Disheveled in Drosophila
Abstract
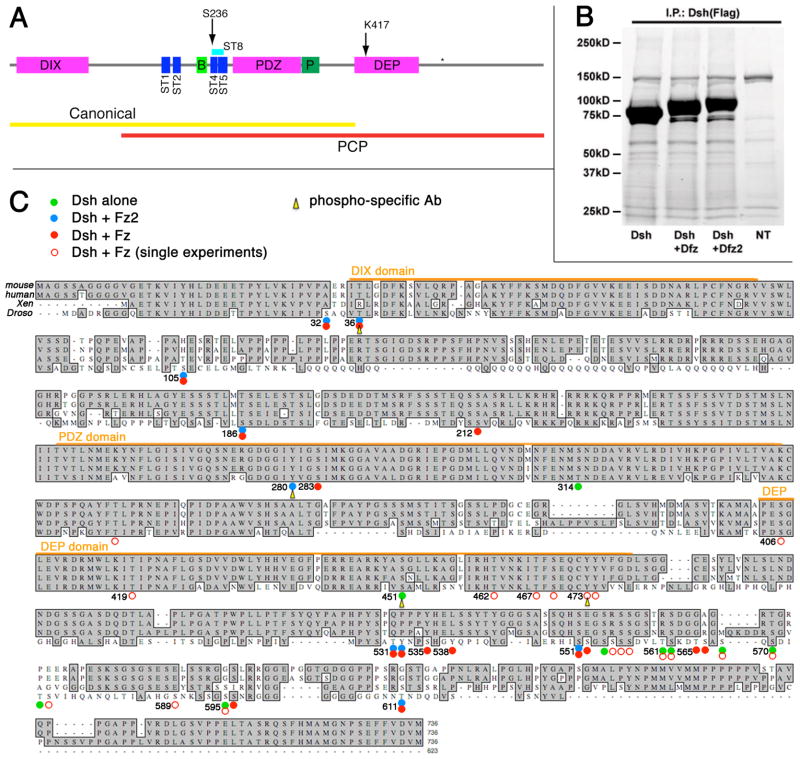
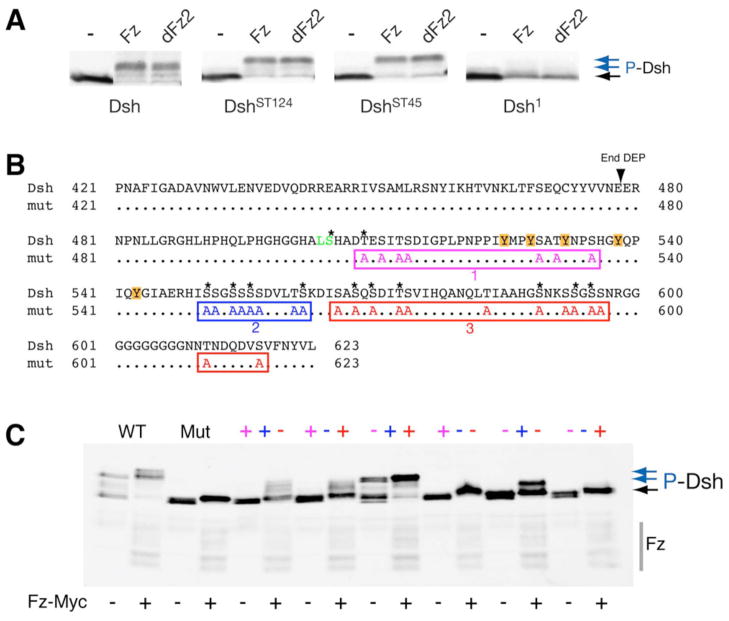
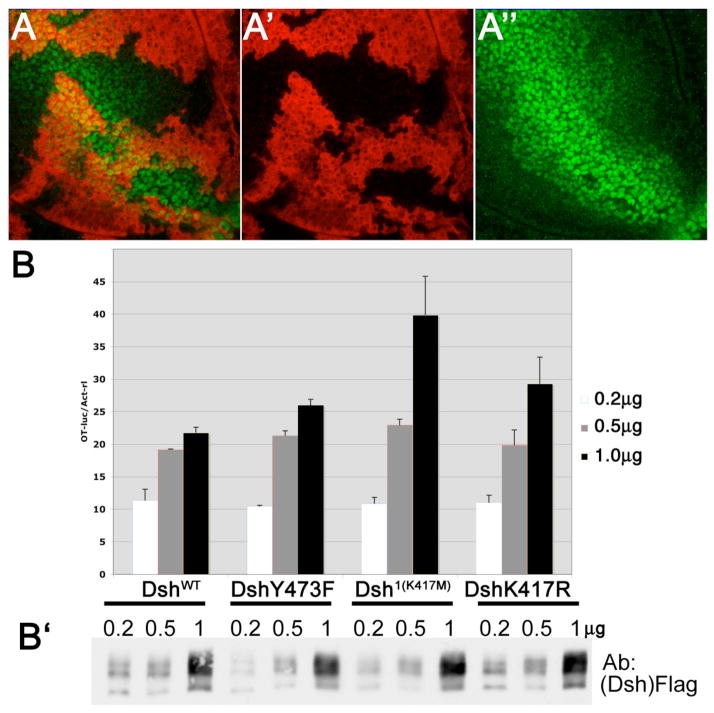
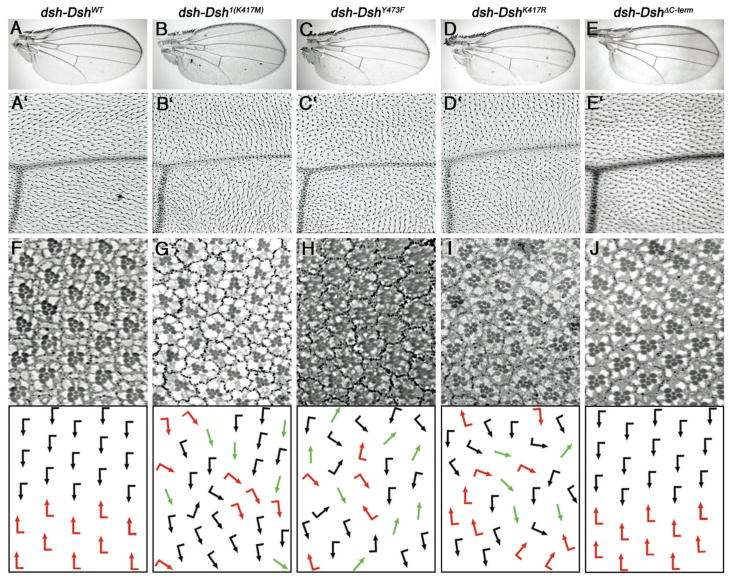
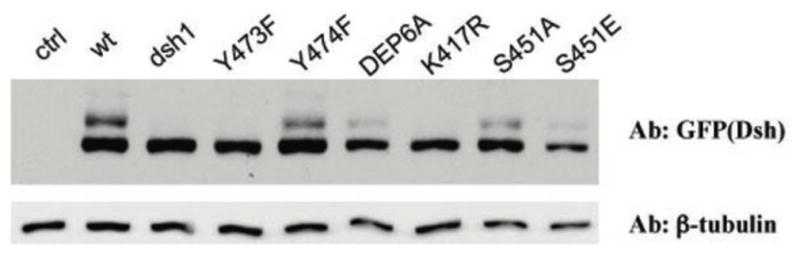
Disheveled/Dsh proteins (Dvl in mammals) are core components of both Wnt/Wg-signaling pathways: canonical β-catenin signaling and Frizzled (Fz)-planar cell polarity (PCP) signaling. Although Dsh is a key cytoplasmic component of both Wnt/Fz-pathways, regulation of its signaling specificity is not well understood. Dsh is phosphorylated, but the functional significance of its phosphorylation remains unclear. We have systematically investigated the phosphorylation of Dsh by combining mass-spectrometry analyses, biochemical studies, and in vivo genetic methods in Drosophila. Our approaches identified multiple phospho-residues of Dsh in vivo. Our data define three novel and unexpected conclusions: (1) strikingly and in contrast to common assumptions, all conserved serines/threonines are non-essential for Dsh function in either pathway; (2) phosphorylation of conserved Tyrosine473 in the DEP domain is critical for PCP-signaling - Dsh(Y473F) behaves like a PCP-specific allele; and (3) defects associated with the PCP specific dsh(1) allele, Dsh(K417M), located within a putative Protein Kinase C consensus site, are likely due to a post-translational modification requirement of Lys417, rather than phosphorylation nearby. In summary, our combined data indicate that while many Ser/Thr and Tyr residues are indeed phosphorylated in vivo, strikingly most of these phosphorylation events are not critical for Dsh function with the exception of DshY473.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


Similar articles
-
CKIepsilon/discs overgrown promotes both Wnt-Fz/beta-catenin and Fz/PCP signaling in Drosophila.Curr Biol. 2006 Jul 11;16(13):1337-43. doi: 10.1016/j.cub.2006.06.030. Curr Biol. 2006. PMID: 16824922
-
Abelson family kinases regulate Frizzled planar cell polarity signaling via Dsh phosphorylation.Genes Dev. 2010 Oct 1;24(19):2157-68. doi: 10.1101/gad.1961010. Epub 2010 Sep 13. Genes Dev. 2010. PMID: 20837657 Free PMC article.
-
Frizzled-Dishevelled signaling specificity outcome can be modulated by Diego in Drosophila.Mech Dev. 2008 Jan-Feb;125(1-2):30-42. doi: 10.1016/j.mod.2007.10.006. Epub 2007 Oct 17. Mech Dev. 2008. PMID: 18065209 Free PMC article.
-
Dishevelled: The hub of Wnt signaling.Cell Signal. 2010 May;22(5):717-27. doi: 10.1016/j.cellsig.2009.11.021. Epub 2009 Dec 13. Cell Signal. 2010. PMID: 20006983 Review.
-
Planar polarity from flies to vertebrates.J Cell Sci. 2004 Feb 1;117(Pt 4):527-33. doi: 10.1242/jcs.00973. J Cell Sci. 2004. PMID: 14730010 Review.
Cited by
-
Autoinhibition of Dishevelled protein regulated by its extreme C terminus plays a distinct role in Wnt/β-catenin and Wnt/planar cell polarity (PCP) signaling pathways.J Biol Chem. 2017 Apr 7;292(14):5898-5908. doi: 10.1074/jbc.M116.772509. Epub 2017 Feb 21. J Biol Chem. 2017. PMID: 28223363 Free PMC article.
-
Regulation of Dishevelled DEP domain swapping by conserved phosphorylation sites.Proc Natl Acad Sci U S A. 2021 Jun 29;118(26):e2103258118. doi: 10.1073/pnas.2103258118. Proc Natl Acad Sci U S A. 2021. PMID: 34155117 Free PMC article.
-
The Wnt/β-catenin pathway in human fibrotic-like diseases and its eligibility as a therapeutic target.Mol Cell Ther. 2015 Jan 30;3:1. doi: 10.1186/s40591-015-0038-2. eCollection 2015. Mol Cell Ther. 2015. PMID: 26056602 Free PMC article.
-
The cellular story of dishevelleds.Croat Med J. 2014 Oct;55(5):459-67. doi: 10.3325/cmj.2014.55.459. Croat Med J. 2014. PMID: 25358879 Free PMC article. Review.
-
WNK Kinases in Development and Disease.Curr Top Dev Biol. 2017;123:1-47. doi: 10.1016/bs.ctdb.2016.08.004. Epub 2016 Sep 28. Curr Top Dev Biol. 2017. PMID: 28236964 Free PMC article. Review.
References
-
- Adler PN. Planar signaling and morphogenesis in Drosophila. Dev Cell. 2002;2:525–535. - PubMed
-
- Bhanot P, Fish M, Jemison JA, Nusse R, Nathans J, Cadigan KM. Frizzled and DFrizzled-2 function as redundant receptors for wingless during drosophila embryonic development [In Process Citation] Development. 1999;126:4175–86. - PubMed
-
- Bhat KM. Frizzled and Frizzled2 play a partially redundant role in Wingless signaling and have similar requirements to Wingless in neurogenesis. Cell. 1998;95:1027–1036. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
